G Protein-Coupled Receptor Family
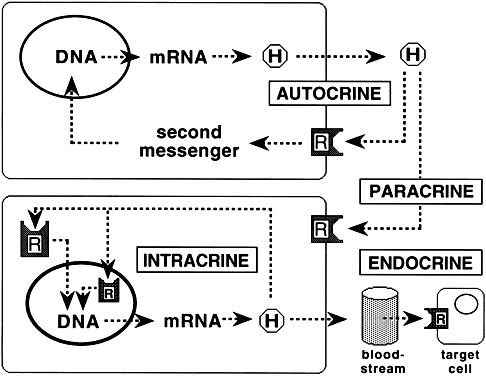
Molecular cloning studies and more traditional biochemical and pharmacologic approaches have revealed a large superfamily of seven transmembrane-spanning receptor proteins that exhibit considerable overlap in their structural features and biologic activities.2,15,16,17,18,19 The receptors bind to G proteins that couple them to a variety of effectors that mediate target cell responses20 (Figs. 1 and 2). The GPCR superfamily can be divided into three groups on the basis of structure:
|
|
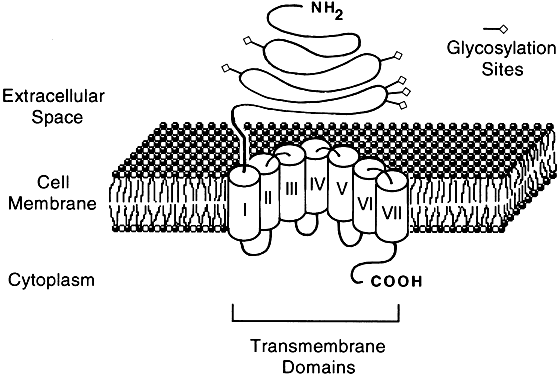
- Group I, composed of receptors that contain 300 to 400 amino acids, has
a short N-terminal extracellular region and binds small ligands (<2 kd), such
as hypothalamic-releasing hormone and α- and β-adrenergic
agents.
- Group II (Fig. 2), which shares considerable homology to group I in the transmembrane region, is
composed of receptors that are 700 to 800 amino acids long, contains
a large extracellular domain, and binds large ligands (30 to 40 kd), such
as the glycoprotein hormones.
- Group III receptors, which share very little homology within the seven
transmembrane regions with the other two groups, are 400 to 600 amino
acids long, have a large extracellular domain, and bind ligands of intermediate
size (4 to 10 kd), such as calcitonin and vasoactive intestinal
peptide.21
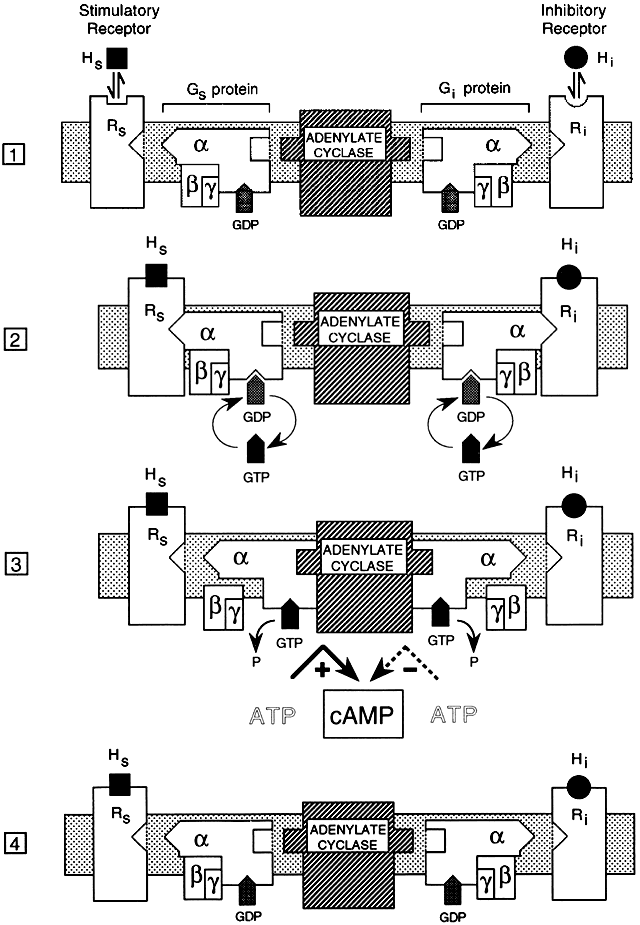
G proteins that couple hormone receptors to effector systems are only one class of a large group of intracellular guanosine triphosphate (GTP)-binding regulatory proteins. There are morethan 1000 members of the GPCR family.22 GPCRs in this class are derived from at least 16 different genes,20 contain α, β, and γ subunits, and have high molecular weights.23 This group of G proteins can endow the hormone receptor with stimulatory (Gs) or inhibitory (Gi) effects on intracellular second messengers, depending on the type of α subunit contained within the G proteins. Gs proteins activate adenylyl cyclase, and Gi proteins inhibit adenylyl cyclase.18 Certain ligands have stimulatory and inhibitory receptor forms, most notably adrenergic receptors, which can activate (β-adrenergic) or inhibit (α2-adrenergic) adenylate cyclase.5
All signal-transducing GTP-binding proteins operate as molecular switches by sequentially binding and hydrolyzing GTP.23 For the detailed mechanism, see Figure 1. Binding of a low-molecular-weight agonist to sites within the hydrophobic core formed by transmembrane helices of the receptor induces dimerization and activation by inducing conformational changes in the receptor protein.24 Once activated by G protein binding, adenylate cyclase catalyzes the formation of cAMP from ATP, which leads to activation of protein kinase A, which phosphorylates other cellular proteins, initiating a cascade of other cellular events.
The α subunit has multiple functional domains that are involved in receptor binding, GTP binding, GTP hydrolysis, α- and β-subunit association, and effector interaction and regulation.25 The βγ complex has been proposed to play a structural role in anchoring the G protein to the cell membrane.26,27 Data indicate that the βγ dimer may also play an important functional role in signal transduction.28 G protein regulation of second messenger activity is complex, exhibiting multiple mechanisms and considerable tissue and receptor specificity. Because each of the subunits exists in multiple forms, there are many possible combinations of the three subunits that comprise the whole G protein complex, each with potentially different activities.
Although G proteins were first described in their role as regulators of adenylate cyclase activity, they also modulate the activity of other second messenger systems and ion channels. Each G protein has unique binding kinetics and affinities for various constituents of the signal transduction cycle.29,30,31 The slow intrinsic rate of GTP hydrolysis by Gα proteins is regulated by interactions with GTPase-activating proteins (GAPs).32 There is a newly discovered family of at least 19 mammalian genes for GAPs known as regulators of G protein signaling (RGS) proteins.32 These proteins interact with various cellular constituents involved in feedback regulation of G protein function and coordination of G protein-regulated signaling systems.32
The mechanisms by which GPCRs lead to cell proliferation is beginning to be understood. In brief, GPCRs and tyrosine kinase receptors activate RAS, thereby initiating a cascade of events leading to activation of mitogen-activated protein (MAP) kinases.33 MAP kinases are critical components of growth-promoting pathways and act by phosphorylating key enzymes and nuclear proteins that ultimately regulate the expression of genes essential for proliferation.22 A novel family of enzymes closely related to MAP kinase, designated JUN kinases (JNKs), selectively regulate the activity of the JUN protein, which is a leucine-zipper family transcription factor.22
Glycoprotein Hormone Receptor Subfamily
The gonadotropins, luteinizing hormone (LH), human chorionic gonadotropin (hCG), folliclestimulating hormone (FSH), and thyrotropin (TSH) are composed of an identical α subunit and a unique β subunit. LH and hCG have a high sequence homology in the β subunit and share a common receptor protein. All three receptors have been cloned and have an overall 55% sequence homology.34 In addition to having seven transmembrane domains, all glycoprotein hormone receptors have a large extracellular N-terminal region and a short cytoplasmic C-terminal domain (Fig. 2). The only region in the glycoprotein hormone receptors that is homologous with other GPCRs is the transmembrane domain.35 The large extracellular domain in the LH/hCG receptor is necessary and sufficient for binding hCG.36
Catalytic Receptor Family
Effector activity is an intrinsic part of the receptor peptide structure for the catalytic receptor family, a group of cell membrane receptors (Fig. 3). Once activated by ligands,5 these single-pass transmembrane proteins act directly as phosphorylating enzymes, mostly through a cytoplasmic domain that functions as a tyrosine-specific protein kinase. Some hormone receptors, such as those for transforming growth factor-β (TGF-β) and for the gonadal peptides, activin and inhibin, act as serine-specific kinases. Threonine-specific receptor kinases have also been described.2
|
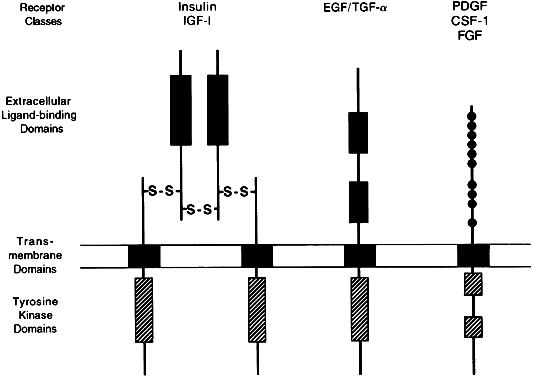
Molecular cloning studies have described three different classes of receptors that contain tyrosine-kinase activity. The most complex class includes receptors for insulin and insulin-like growth factor-1 (IGF-1), with two α and two β subunits, forming two transmembrane domains connected extracellularly by disulfide bridges.37 The second class, represented by the epidermal growth factor (EGF)/TGF-β receptor, contains an extracellular growth factor-binding domain connected to a cytoplasmic tyrosine kinase domain by a single transmembrane region.38 The third class includes receptors for plateletderived growth factor, fibroblast growth factor, and colony-stimulating factor-1, which contain a single transmembrane domain and a cysteine-rich extracellular domain.39,40
Autophosphorylation of receptors in tyrosine, serine, or threonine residues by the kinase portion of receptor protein or receptor phosphorylation by other cellular kinases is thought to be an important mechanism of receptor activation,41,42,43 possibly by affecting the tertiary structure of the receptor protein.37 Autophosphorylation is also an effective positive feedback mechanism, providing signal amplification. Cross-phosphorylation of receptors for other hormones, such as the phosphorylation of IGF-1 receptors by insulin receptors, may contribute to the diversity of effects of these hormones in target cells.44
Although only a few of the specific protein substrates for receptor tyrosine kinases have been identified, some phosphoproteins are common substrates to several receptors, suggesting another mechanism for overlap in cellular activities of certain hormones.45 In addition to tyrosine phosphorylation, insulin, EGF, and other growth factors appear to regulate cell function through control of other second messenger systems. For example, phosphoinositol turnover ultimately results in activation of protein kinase C and changes in Ca2+ flux46 (Table 1). In the case of EGF, there is evidence for cAMP playing a second messenger role. IGF-1 was reported to activate a nuclear phosphoinositide signaling system that is entirely separate from membrane-bound receptor signaling systems.47
Cloning and characterization of the growth hormone48 and prolactin49 receptors revealed that both are closely related single-pass transmembrane proteins, with structural similarities to cell transport molecules (transferrin) and some homology with erythropoietin, CSF-1, and several interleukins.49 Receptors for growth hormone and prolactin have also been reported to contain or be associated with tyrosine kinase activity, although it is unclear whether tyrosine kinase is an intrinsic property of the receptor molecule.50 Prolactin and growth hormone stimulate protein kinase C activity in vitro51 and appear to phosphorylate the same high-molecular-weight protein.
Channel-Linked Receptor Family
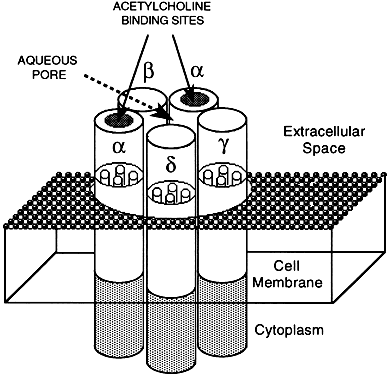
Hormone receptors in this group function as ligand-gated ion channels and belong to a family of homologous, multipass transmembrane proteins (Fig. 4). Channel-linked receptors regulate the flux of calcium, potassium, chloride, and sodium ions across the cell membrane. Ion channels allow movement of ions at very rapid rates (107 to 109 ions per second) across membranes, several orders of magnitude higher than most active transport mechanisms.52 Ion channel-linked receptors are involved predominantly in rapid chemical synaptic neurotransmission between electrically excitable cells and may also provide a mechanism for autocrine or modulatory effects of certain hormones and neurotransmitters.7 The best understood ligand-gated ion channel is the directly channel-coupled nicotinic cholinergic receptor, consisting of five glycosylated subunits, encoded by separate genes, with each subunit containing four transmembrane-spanning domains53 (Fig. 4). Two major types of ion channel modulation involve indirect, longer-duration mechanisms: phosphorylation by protein kinases and interaction with G proteins. Cyclic AMP-dependent kinases and other kinases phosphorylate the nicotinic acetylcholine receptor, affecting the rate of desensitization and modulating ion channel activity.54,55 Noncovalent interactions between G proteins and membrane ion channel proteins, such as the muscarinic acetylcholine-regulated potassium channel and the β-adrenergic-gated calcium channel,56 also appear to play an important role in ion channel gating.2,57
|