Human Papillomavirus Infection and Genital Tract Cancer
Authors
INTRODUCTION
During the past 20 years, a large number of studies have shown a strong association between the presence of specific types of human papillomavirus (HPV) and the development of anogenital cancer. It is estimated that infection with HPV is globally associated with 561,100 cancers in women and men, ranging from cancer of the cervix, vulva, vagina, anus, and penis, to that of the mouth and oropharynx.1 Cervical cancer is of particular importance among anogenital cancers because it is the most common cancer in women, with 493,000 new cases diagnosed each year and 274,000 deaths documented in 2002.2 DNA of specific HPV types has been found outside the anogenital sphere in some squamous cell carcinomas of the skin, tongue, oral cavity, hypopharynx, larynx, and lung.3
HISTORICAL PERSPECTIVE
Early evidence
Isolated cases of conversion of genital warts into squamous cell carcinomas of the vulva and penis were first reported in the 19th century. The formation of the neoplasms was not interpreted as being related to an infectious event until Shope, in 1933, demonstrated the infectious nature of warts by infecting rabbits with papilloma extracts from wild cottontail rabbits.4 The cottontail rabbit papillomavirus was later named after Shope and was shown to induce rabbit papillomas that developed into malignant squamous cell carcinomas when exposed to chemical mutagens and radiation.2 Because it was, until recently, impossible to grow HPV in tissue culture or to transfer it to suitable animal hosts, further progress was slow until advances were made in molecular biology with hybridization techniques and molecular cloning.
Discovery of nonpapillomatous HPV-associated genital lesions
In 1976, zur Hausen postulated that HPV played a role in anogenital carcinoma. In the same year, Meisels and Fortin3 and almost simultaneously Purola and Savia5 described epithelial changes with cytologic features identical to those of condyloma acuminatum, but without their papillary appearance. The lesions were clinically indistinguishable from low-grade cervical dysplasia and were called flat condyloma. It also was apparent that these subclinical epithelial changes were much more common than papillomatous forms of the infection. The cells in the flat condylomas were called koilocytes. The term (from the Greek koilos, meaning hollow) reflects the hollow appearance of the cells and was originally coined by Koss and Durfee in 19566 to describe cells having perinuclear cytoplasmic clearing, or “halos”, in cytologic specimens from both cervical cancers and cancer precursors. Because condylomata acuminata were known to be of viral origin, the demonstration of koilocytes in both genital warts and dysplasia suggested that cervical neoplasias also have a viral cause.
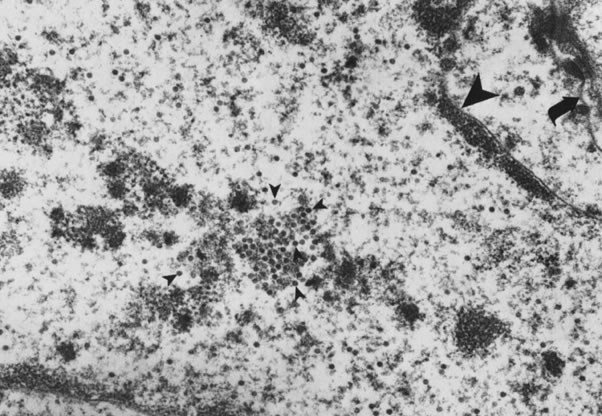
In 1978, Della Torre and coworkers7 and Laverty and associates8 detected viral particles in flat condylomata using electron microscopy (Fig. 1). Direct evidence that the virus was actually HPV came after Jenson and coworkers9 developed group-specific antibodies acting against capsid proteins of a wide variety of animal and human papillomaviruses. The immunoperoxidase technique employing these anti-HPV antibodies demonstrated papillomavirus in the superficial layers and in koilocytotic cells of about half of flat condylomata and mild dysplasias.10 The demonstration of HPV DNA in cervical cancer had to await the application of molecular probes in the early 1980s. By that time, it was generally accepted that the koilocytotic change is the cytomorphologic hallmark of the cytopathic effect of HPV on the superficial epithelial cells, and that condylomata and dysplasia are forms of expression of HPV infection.11
|
EPIDEMIOLOGY OF HPV INFECTION
Prevalence
Genital HPV infection is the most common sexually transmitted infection in women. Among asymptomatic women in the general population, the prevalence of HPV infection ranges from 2 to 44%.12 A recent meta-analysis designed to estimate the age and genotype specific prevalence of cervical HPV DNA in women with normal cytology, systematically reviewed 78 studies published between 1995 and 2005.13 The analysis showed that overall 10.4% (95% CI 10.2–10.7) of over 150,000 women with normal cytology were HPV DNA positive. There was some regional variation with HPV prevalence in Africa reaching 22.1%, Central America and Mexico 20.4%, northern America 11.3%, Europe 8.1%, and Asia 8.0%. Many studies have shown an age-related decline in HPV prevalence, even in a study of prostitutes (who are highly exposed to HPV) Kjaer et al. showed a decrease in HPV prevalence with age despite high levels of sexual activity.14 It has been postulated that this occurs because infected individuals develop adaptive immune responses against HPV that prevent future infection.
Using the polymerase chain reaction (PCR), Bauer and coworkers15 found that 46% of 467 women presenting to a university health service for a routine annual gynecologic examination tested positive for HPV. Sixty-nine per cent of the HPV-infected women tested positive at both sites examined, cervix and vulva, indicating that HPV infections involve the entire lower genital tract. Thirty per cent of the study population harbored the potentially oncogenic HPV types. The high prevalence of latent infections may define a large risk group from which active infection and neoplasia may emerge. HPV-infected women, however, often produce immune responses that eventually clear the virus. For this reason, women aged 35–55 years participating in cervical cancer screening programs were less often positive for HPV than sexually active women between 15 and 30 years of age.16, 17 These data are confirmed by the meta-analysis of Sanjose et al.17 Only the number of sexual partners during recent years (1–5 years) appears to be associated with positive HPV tests.18
Incidence of HPV infection
Studies that have repeatedly measured the presence of HPV DNA over time show that the cumulative risk of acquiring HPV in sexually active women and men is high. For example, among women aged 15–19 years in England, and among two cohorts of college women in the USA, the cumulative incidence of HPV infection was more than 40% over 3 years.19, 20, 21 In the Brazilian Ludwig-McGill cohort with women having a mean age of 33.3, the cumulative incidence was 24% over 18 months.22
Persistence of HPV infection
HPV infections occur very commonly, but most infections clear spontaneously and only a small proportion of infections persist over time.23, 24 Whether infections clear completely or the virus remains latent in basal cells at undetectable levels is unclear and has not been empirically verified. What has been shown, however, is that the risk of subsequent cervical intraepithelial neoplasia (CIN) is significantly increased by persistent HPV infection over long periods of time, particularly with infection by high-risk types of HPV.25
Transmission
The most common mode of transmission is by sexual activity through contact with infected cervical, vaginal, vulvar, penile, and anal epithelium. Sexual contact with an infected partner is necessary for transmission and it is thought that this occurs through microscopic abrasions in the mucosa or skin. "Skin to skin" contact is essential but intromissive intercourse is not strictly necessary. HPV infections can be transmitted by peno–vaginal intercourse and by other sexual practices such as oral sex, peno–anal intercourse, digital–vaginal sex, and use of insertive sex toys.26, 27 Receptive anal sex is strongly associated with HPV detection in the anal canal in homosexual and bisexual men, and to a lesser extent in women.28 Non-sexual modes of transmission of genital HPV remains a controversial issue. Non-sexual modes of transmission include perinatal transmission, and fomites and skin contact (auto-innoculation). While these routes are possible they are extremely rare.
Genital condylomata acuminata are highly contagious, with an infectivity rate of at least 60%. HPV-6 and to a lesser extent HPV-11 are the types most readily transmitted, probably because exophytic and often friable lesions caused by these HPV types release a large amount of infectious material. In addition, condylomata acuminata contain a much higher number of viruses per infected cell than other HPV-associated lesions.29 Although these factors favor dissemination of HPV-6 and HPV-11, the viruses are uncommon in healthy women. Only 3% of asymptomatic women undergoing a routine annual gynecologic examination had HPV-6 or HPV-11, whereas 43% harbored other HPV types.15
Despite a lower viral load, subclinical condylomata and intraepithelial neoplasia associated with HPV types other than 6 and 11 appear to be as infectious as condylomata acuminata. About 65% of male partners of women with subclinical lesions detected by cervical cytologic smears have HPV-associated penile lesions.30
Although the major route of transmission of genital HPV is by sexual contact, perinatal transmission also is possible and has been documented in 55% of infants passing through an infected birth canal. Persistent HPV infection was demonstrated in some of the infants. Whether acquisition of HPV during the perinatal period predisposes to an increased risk of genital carcinoma later in life remains to be established.31 Perinatal transmission of HPV has been demonstrated unequivocally for the rare disease juvenile respiratory papillomatosis.32
Incubation
The incubation period ranges from 3 weeks to months, sometimes longer, the average being 2.8 months. In general, across many studies averages for individual infectious episodes range from 4 to 20 months. Clearance rates vary considerably but range from 40 to 90%. Further, infections with low-risk types of HPV last for shorter times than high-risk types of HPV.
The incubation period from the time of infection to the establishment of noncondylomatous HPV infections is unknown but may be similar to that of papilloma-forming viruses. Dysplasia may develop primarily within latently infected epithelium or secondarily from established subclinical lesions. Follow-up studies of women who tested positive for HPV DNA indicate a lag period of 6–24 months before a cytologic abnormality occurs.16 The transformation from viral infection to carcinoma most often takes several years, sometimes decades.
HPV infection as the causal agent of cervical cancer
It is now well established that infection with high-risk types of HPV (HR-HPV) is a necessary for, albeit not sufficient cause of, almost all cervical cancers.33 In the study of Walboomers et al. successive PCR analyses of a large international collection of cervical tumor specimens, showed that HPV DNA was present in 99.7% of specimens. Relative risks for the association between HPV and cervical cancer are in the 50–150 range.
Risk factors for HPV infection have been identified in a range of cross-sectional and prospective cohort studies and these include number of sexual partners (lifetime and recent), age at first intercourse, smoking, oral contraception, other STIs (chlamydia and herpes simplex virus), chronic inflammation, immunosuppressive conditions including HIV, and parity.34, 35, 36 In addition to the correlates of sexual activity, the most important risk factor for HPV infection is age, with most studies indicating a sharp decrease in infection rates after the age of 30 years.
CLASSIFICATION OF PAPILLOMAVIRUSES
Papillomaviruses belong to the family Papovaviridiae, which includes polyoma viruses and SV40 viruses. They are all double-stranded DNA tumor viruses but lack similarities in DNA sequence homology, size, and antigenicity. These epitheliotropic viruses infect the surface epithelia and mucous membranes, where they produce warts and epithelial proliferations. They are found in a wide variety of vertebrates and are highly species-specific. The classification of papillomaviruses is based on the species of origin and the extent of relatedness between viral genomes. Within a given species, there may be a large number of different types of papillomavirus, referred to as genotypes rather than serotypes because they are classified on the basis of DNA composition as opposed to antigenicity. The viruses are assigned numbers according to their order of discovery. It was originally agreed to designate a new papillomavirus as a different type if its DNA is less than 50% homologous to DNA of other known types of papillomavirus from the same species.37 More recently, HPV types are classified as distinct if the E6, E7, and L1 gene sequences differ by more than 10% from those of other known HPV types. In addition to types, there are also subtypes or variants of specific types. To be considered a subtype or variant, the different viruses must differ by 2–5% from the original isolate.38
More than 120 genotypes of HPV have been characterized. Thirty-three of the HPV types occur predominantly or exclusively in the anogenital tract (Table 1). HPV-16 and HPV-18 are most commonly encountered in anogenital cancers, being present in about 60–80% of biopsy samples. Many of the other 33 types have been found only sporadically in individual tumors.39, 40
More recently, genital HPV types have been subdivided into low-risk types (LR) which are found mainly in genital warts and low grade intraepithelial abnormalities, and high-risk types (HR), which are frequently associated with cervical cancer. Munoz et al.41 conducted 11 case–control studies in nine countries between 1985 and 1997 and performed HPV DNA typing using polymerase chain reaction (PCR)-based assays. A total of 1990 of 2365 women with squamous cell carcinoma and 2126 of 2491 control women provided cervical specimens for HPV DNA testing. HPV DNA was detected in 96.6% of the women with cervical cancer and 15.6% of the control women. The pooled odds ratio for cervical cancer associated with the presence of any HPV was 158.2 (95% CI 113.4–220.6). On the basis of this study, 15 HPV types were classified as HR-types: 16, 18, 31, 33, 35, 39, 45, 51, 52, 56, 58, 59, 68, 73, and 82. Three were classified as 'probably HR-types': 25, 53, and 66. A further 12 were classified a low-risk types: 6, 11, 40, 42, 43, 44, 54, 61, 70, 72, 81, and CP6108.
Table 1. HPV types and their association with diseases
HPV Type | Associated Disease |
1, 2, 4 | Plantar and common warts |
3, 28, 29 | Flat warts |
7 | Common warts of meat and animal handlers |
5, 8, 9, 10, 12, 14, 15, 17, 19, 20, 21, 22, 23, 24, 25, 36, 37, 38, 47, 50 | Flat warts, macules and pityiasis, versicolor-like lesions in epidermodysplasia verruciformis (EV) patients, some types occasionally found in keratoacanthoma, solar keratosis, and melanoma |
6, 11, 42, 43, 44, 45, 54, 55, 57 | Anogenital condylomata acuminata |
6, 11 | Laryngeal and conjunctival papilloma |
6, 11, 16, 18, 30, 31, 33, 34, 35, 39, 40, 41, 42, 43, 44, 45, 51, 52, 53, 54, 55, 56, 57, 58, 59, 61, 62, 64, 66, 67, 68, 69, 70 | Cervical intraepithelial neoplasia (CIN), vaginal intraepithelial neoplasia (VAIN), penile intraepithelial neoplasia (PIN), vulvar intraepithelial neoplasia (VIN) |
13, 32 | Oral focal epithelial hyperplasia (Heck's disease) |
26, 27, 49 | Cutaneous warts from a patient with immune deficiency and from a renal transplant recipient |
41, 48 | Disseminated warts, squamous cell carcinoma of the skin |
46 | EV-like lesions in a patient with Hodgkin's disease |
53 | No specific disease (cloned from cervical scraping) |
(Modified from Dürst M: The human papillomaviruses: Classification and molecular biology. Clin Prac Gynecol 2: 29, 1989)
DETECTION OF HPV
Culture
Limited quantities of complete HPV can now be produced in transplants of human tissues in nude mice. The tissues such as neonatal foreskin, cervix, or larynx are exposed to extracts of condylomas containing HPV and are then transplanted beneath the renal capsule of nude mice. After several months, cytopathic effects and capsid proteins from the virus can be detected in the transplanted human tissue.42 Another method of propagating HPV is the collagen-raft culture system. This method involves culturing human keratinocytes infected with HPV in organotropic (i.e., collagen-raft) media. Chemical treatment stimulates differentiation of the keratinocytes and, simultaneously, biosynthesis of complete HPV.43 Unfortunately, all known culture methods are cumbersome and limited to few HPV types and thus are not suitable for clinical or epidemiologic studies.
Immunologic methods
Because adequate amounts of complete human papilloma virions are not yet obtainable through culture or clinical lesions, it has not been possible to date to identify appropriate antigens for immunologic assays. To circumvent this problem, HPV-specific proteins produced in bacteria were used to search for antibodies against HPV. The detected antibodies against HPV E7 proteins were found in about half of women with cervical cancer but not at all in women with precursor lesions or subclinical HPV infections.44 Therefore, antibodies against E7 proteins are not suitable to screen women for exposure to high-risk HPV types.
A different approach to developing specific serologic markers for HPV infection consists of assembly of HPV capsids in insect cells exposed to HPV L1 protein. The L1 protein encodes the development of the viral capsid but not the viral DNA core. Specific antibodies against the “empty” capsid or viral particle have been found in 59% of women with HPV-16 and in 92% of women with condylomata acuminata.45 Virus-like particles are being studied as substrates for HPV vaccines and are expected to be of major importance in the future.38
DNA typing
The isolation and characterization of individual HPV types was hampered by the low particle production in HPV-associated lesions. Fortuitously, Gissmann and zur Hausen40 encountered a rare condyloma with high particle yield that permitted partial characterization of viral DNA in 1980. The DNA from this lesion was cloned in bacterial vectors and resulted in sufficient quantities of viral DNA for further characterization of the virus that was later labeled HPV-6. Subsequently, the known DNA of HPV-6 was labeled with a radioactive tracer and was used as a probe in hybridization procedures for identification of related but distinct viral types.
Shortly after discovery of HPV-6, the hybridization technique permitted isolation of the closely related HPV-11 from laryngeal papillomas and genital warts. In addition, DNA of the only distantly related virus types, HPV-16 and HPV-18, was directly cloned from cervical cancer biopsies.46, 47
HPV DNA tests used in clinical and epidemiological trials
Currently, HPV DNA tests that have been validated in large clinical and epidemiological trials are the second generation hybrid capture (HC2) test and PCR-based methods employing either MY09/11 or GP5/6 consensus primers. The hybrid capture HPV DNA assay is based on hybridization in solution of long synthetic RNA probes complementary to the genomic sequence of 13 HR-types (16, 18, 31, 33, 35, 39, 45, 51, 52, 56, 58, 59, and 68). DNA present in the specimen is hybridized in solution allowing the formation of specific HPV DNA–RNA hybrids. These hybrids are then captured by antibodies bound to the wells of a microtiter plate that recognize specific RNA–DNA hybrids. After removal of excess antibodies and non-hybridized probes, the immobilized hybrids are detected by a series of reactions that give rise to a luminescent product which is measured by a luminometer. The intensity of emitted light, expressed as relative light units (RLUs) is proportional to the amount of target DNA present in the specimen.48
PCR-based assays
PCR-based methods have the highest analytical sensitivity and can detect as few as 10–100 copies of HPV genomes in the specimen being tested. In addition, PCR-based assays require very small amounts of specimen. The assays are designed to selectively amplify the viral genome by a series of polymerization steps, which result in an exponential and reproducible increase in the nucleic acid sequences present in the biological specimen. The assays rely on the amplification of selected portions of the gene of interest the boundaries of which are defined by oligonucleotides that hybridize (anneal) to their complementary sequences on the target strand that has been previously denatured. At defined temperatures, such oligonucleotides are extended by a thermoresistant DNA polymerase leading to the formation of two new double-stranded DNA molecules (known as the amplicon) using each of the original target DNA single strands as templates. By repeating the cycle of denaturation, annealing, and extension, each newly synthesized double-stranded DNA molecule can serve as a template for the next cycle, and the number of molecules increases in an exponential fashion. Analysis of the amplified products may be performed by gel electrophoresis, dot blot or line strip hybridization, restriction length polymorphism analysis or sequencing.49
METHODS OF HPV IDENTIFICATION
Hybridization technique
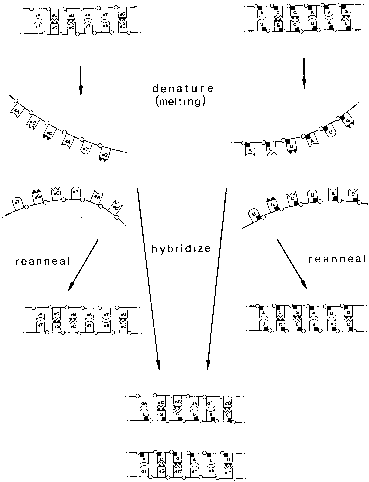
The basic principle of the hybridization technique is the duplex formation of single-stranded DNA (or RNA) derived from the cloned HPV probe and the viral DNA target molecule in the cell or tissue to be examined. After dissociation of the target DNA into single DNA strands through denaturization or melting, the more-or-less complementary single DNA strands of probe and target are allowed to associate. The resulting probe–target complexes show different stability, which is directly related to the degree of homology between the probe and target molecules and the experimental condition. Through variations of temperature or chemical parameters, different levels of stringency can be created. For example, under conditions of low stringency, the probe binds to target DNA of high and low homology. It is, therefore, suitable for screening for related or new HPV types. Conditions of high stringency, by contrast, identify only closely related or identical HPV DNA because DNA with low homology dissociates from the probe (Fig. 2).
|
Filter hybridization
Among currently used methods of HPV-DNA identification, filter hybridization techniques, such as Southern blot, dot-blot, and filter in situ hybridization (FISH), play a less important role. For these tests, nitrocellulose filters provide the base to which viral DNA and probe DNA are applied. In the Southern blot method, the target DNA is cut with restriction enzymes at specific sites, fractionated according to size by gel electrophoresis, transferred to the filter, and then hybridized with radiolabeled HPV probes50 (Fig. 3).
|
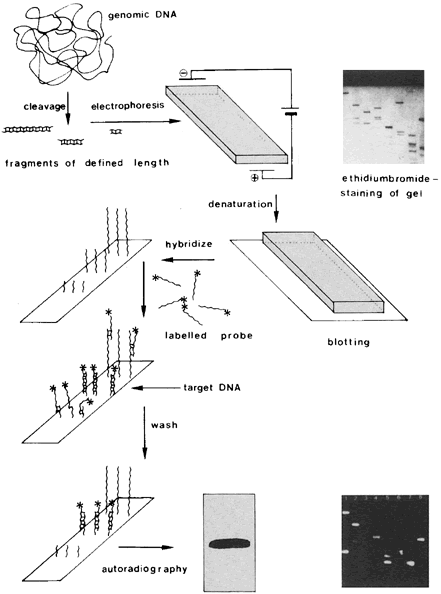
The dot-blot or slot-blot analysis is a variant of the Southern blot method in which the extracted target DNA is applied directly to the filter in either a dot or slot pattern and then hybridized with the labeled probe. The FISH method is a simplification of the dot-blot analysis process and does not require prior DNA extraction. Instead, cytologic or biopsy specimens are placed in a lysis buffer; the DNA in the lysate is then denatured, transferred to a filter support, and hybridized with a labeled HPV probe.51 A drawback to this method is that it does not allow quantitation of viral DNA.
The Southern blot analysis is highly sensitive and specific, and permits identification of DNA fragments characteristic of different HPV types. It also allows assessment of the physical state of HPV DNA in the cells (i.e., integrated vs. episomal) and has been regarded as the gold standard of HPV nucleic acid hybridization techniques. The method is time-consuming, however, and therefore not practical for screening of samples on a large scale. HPV testing kits using the FISH technology were the first to be approved by the Food and Drug Administration (FDA) for clinical use and made commercially available. Because of their high cost and low sensitivity and specificity, FISH-based tests did not gain wide popularity and have been replaced by solution hybridization and PCR.
In situ hybridization
Besides the filter hybridization methods, in situ hybridization of tissue sections is important. In this technique, isotope probes or nonradioactive biotin-labeled probes are added directly to cytologic preparations or histologic sections on a glass side. In situ hybridization may be carried out on frozen or fixed materials. It is, therefore, suitable for retrospective studies and allows localization of viral nucleic acid sequences in cytologic or histologic specimens (Fig. 4). Because of its relative insensitivity compared with filter hybridization methods, in situ hybridization is of limited value in the examination of tissues with low cellular concentrations of HPV DNA, such as latently infected epithelia, high-grade dysplasia, and invasive carcinoma.18
|
Polymerase chain reaction
The sensitivity of hybridization techniques may be increased by selective amplification of viral DNA with PCR. In this technique, synthetic oligonucleotide sequences, called primers, bind to specific, homologous sites of separated strands of the target DNA. By adding DNA polymerase to the assay, the primers are extended using the single strands of the target DNA as templates. Through repeated cycles of denaturation, annealing with the oligomer primers, and primer extension with DNA polymerase, the target DNA is amplified exponentially with the number of cycles. It is theoretically possible to detect a single HPV DNA molecule in 1 million cells.52
Its extremely high sensitivity makes PCR an outstanding research tool but may limit its clinical usefulness because traces of HPV DNA may be detected that are clinically meaningless.53 The early PCR methodology often produced false-positive results by amplifying small amounts of contaminating HPV DNA derived from biopsy forceps or laboratory tests. Test kits based on the PCR technology have become commercially available for clinical use.
Solution hybridization
Developments in technology have led to a revival of the oldest nucleic acid hybridization methodology, whereby hybridization occurs in solution. The new versions of the solution hybridization are as sensitive and specific as the Southern blot method but are much simpler to perform.
In the hybrid capture technique, DNA probes are used to bond single-stranded HPV DNA. The hybrids are captured onto the surface of a plastic tube by specific antibodies. The reaction is made visible by the emission of light in direct proportion to the quantity of captured hybrid. A commercial form of the test, Hybrid Capture II (HC 2; Dygene Diagnostics, Silver Spring, MD), has been approved by the FDA. The test detects essentially all HPV types relevant for cervical neoplasia.53 Testing for intermediate- and high-risk HPV by hybrid capture improves the detection of high-grade squamous intraepithelial lesions over cytologic study used alone in women with atypical squamous cells of undetermined significance and low-grade squamous intraepithelial lesions.42, 54 Methods of HPV identification are compared in Table 2.
Table 2. Methods of human papillomavirus identification
Hybridization Method | Sensitivity | Specificity | Labor and Cost | Remarks |
Southern blot | ++ | +++ | +++ | Gold standard |
Dot blot | ++ | ++ | ++ | Used clinically (first-generation tests) |
FISH | + | + | + | Simple, rapid, inaccurate |
In situ | + | + | ++ | For histologic sections |
PCR | +++ | +++ | ++ | Extremely sensitive |
Solution hybridization | ++ | +++ | + | Suitable for clinical use (second generation tests) |
FISH, filter in situ hybridization; PCR, polymerase chain reaction; +, low; ++, moderate; +++, high.
Clinical application of HPV testing
The understanding of the strong causal relationship between HPV infection of the cervix and cervical cancer has prompted a large body of research on the use of HPV DNA testing in cervical cancer prevention programs. Detection of HR-HPV DNA has been suggested in four clinical scenarios: (1) as a primary screening test solely or in combination with cervical cytology;55 (2) as a test to triage women with minor atypical Pap smears to determine who needs to be referred for colposcopic assessment for diagnosis and possible treatment;56 (3) in the management of women referred for colposcopy for whom no lesion is visualized; and (4) as a follow-up test for women treated for cervical cancer precursors.57
HPV DNA testing is without doubt significantly more sensitive than cytology for the detection of cervical cancer precursors, although less specific and with a lower positive predictive value than cytology. In a review of 14 studies comparing cytology with PV DNA testing (both HC 2 and PCR-based) Franco showed that, on average, HPV DNA testing has a 27% higher sensitivity than cytology in absolute terms, with a specificity which is, on average, 8.4% lower than cytology.58 The very high negative predictive value of HPV DNA testing, however, may well enable much longer screening intervals. In a primary screening setting, HPV DNA testing is recommended as an adjunct to cytology in women over the age of 30 years only, due to the very high prevalence of HPV infection in younger women.
The first formally accepted clinical use for HPV DNA testing by the FDA in the USA, was for the triage of atypical squamous cells of unknown significance (ASCUS) cytology. The ALTS trial (ASCUS/LSIL [low-grade squamous intraepithelial lesions] Triage Study) was a large multisite randomized trial specifically designed to evaluate different methods of managing women with ASCUS and LSIL cytology results.59 HPV testing was found to be positive in approximately 80% of women who had a cytological diagnosis of LSIL and the study concluded that HPV testing was not of value in managing women with LSIL cytology. However, the same study found that HPV testing detected 96.3% of women with undiagnosed CIN 3 or cancer who had ASCUS cytology, and that HPV testing resulted in referring 56.1% of women for colposcopy. In settings where the practice is to perform colposcopy on all women with ASCUS cytology, HPV DNA testing would significantly reduce the number of women requiring colposcopy.
Ablative or excisional techniques for the treatment of cervical cancer precursors are generally reported to be very effective, with cure rates of over 90% reported. However, in 5–15% of cases, intraepithelial lesions will either persist or recur. A number of studies57 have shown significantly higher sensitivity and negative predictive value of HPV DNA testing compared to cytology for the detection of recurrence or persistence.
The role of HPV DNA testing may assume increasing importance as liquid-based cytology replaces the conventional Pap smear and “reflex testing” becomes more widely available. With reflex testing, the laboratory is instructed to reflexively perform HPV DNA testing on the residual material in the liquid cytology vial whenever a diagnosis of atypical squamous cells of undetermined significance is made.60
STRUCTURE AND GENOMIC ORGANIZATION OF PAPILLOMAVIRUSES
Papillomaviruses consist of a circular, double-stranded DNA containing about 7800–7900 base pairs, surrounded by an icosahedral capsid. The genomic organization of all papillomaviruses appears to be similar and contains several regions able to code for proteins. These stretches of DNA are known as open reading frames (ORFs), referring to the possibility of reading relatively long segments of genetic code (400 bases or more) before reaching a terminal signal. All of the ORFs are found in one strand of DNA, and therefore all messenger RNA is copied from one strand. Figure 5 shows the arrangement of ORFs for HPV-16.
|
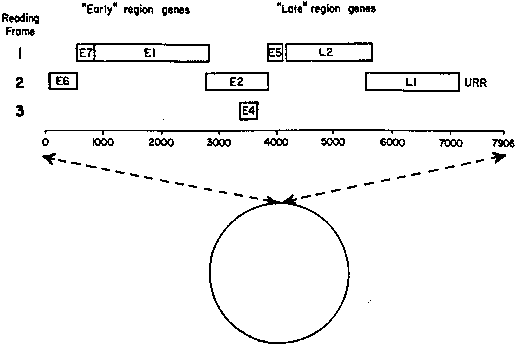
Nine ORFs have been identified among the most known types of HPV. Seven of the ORFs are called early ORFs (E1–E7) because they have important functions related to early events of viral infection, including cell transformation and DNA replication. The other two ORFs follow the early region and are called late ORFs. They are expressed only in the superficial layers of the affected epithelium and code for viral capsid protein production, a late event in the course of viral infection leading to viral assembly. In addition to the coding regions, the viral genome contains a base-pair region located upstream of the open DNA molecule, between the end of the late region and the beginning of the early region. This stretch of the viral DNA controls DNA replication and expression of viral genes and is called the upstream regulatory region.
Transcription of the late region appears to be dependent on signals related to maturation of the host cell. Proteins derived from late ORFs, therefore, are not detected in most cervical cancers. By contrast, proteins coded for by the early ORFs are consistently found in HPV-associated cancers and in cell lines derived from these neoplasms. The E6 and E7 ORFs are of particular importance because they have been implicated as transcribing genes (oncogenes), which activate and propagate the oncogenic process associated with certain HPV types, in particular HPV-16 and HPV-18.
HPV ONCOGENESIS
Immortalization and transformation of cultured cells
Cells from neonatal and adult tissues cultured in vitro replicate only a limited number of times before they stop dividing and die. Cells that continue to replicate indefinitely are designated as being immortalized. Immortalization is a major growth characteristic of cells cultured from malignant neoplasms. HPV-16 and HPV-18 protein coded by the E7 ORF is capable of producing immortalization of cultured cells.61, 62, 63
Another effect of HPV DNA is the capacity to transform already immortalized cells. When HPV-16 or HPV-18 DNA is introduced into an already immortalized cell line, the recipient cells become transformed and develop the ability to grow when suspended in fluid or in a semisolid agar gel. This type of growth is termed anchorage-independent growth and is viewed as a second separate characteristic of malignant cells. Some of these transformed cell lines are also tumorigenic and form tumors in mice. Immortalization and transformation are prerequisites of oncogenesis and are achieved by HPV-16 and HPV-18, but not HPV-6.64
Inactivation of tumor suppressor genes
The most important biologic function of E6 proteins from oncogenic HPV types appears to be inactivation of the p53 tumor suppressor protein. The p53 protein acts as a cellular gate-keeper for growth and cell division by regulating expression of growth-suppressive proteins such as the cell cycle inhibitory protein, p21, and a repressor of p53 itself, mdm-2.65 p53 has been shown to be responsible for mediating cellular growth arrest in response to DNA damage and growth factor deprivation. It has also been shown to regulate programmed cell death, or apoptosis, in cells that have undergone irreparable injury.50
The E6 proteins of oncogenic HPV types consist of approximately 150 amino acids and have sequence homology with both adenovirus E1B protein and SV40 large T antigen. All these proteins are capable of binding to the p53 protein. The result is chromosomal instability, an elevated mutation rate, and progression to malignancy.51 High-risk HPV types bind to the p53 protein with much greater efficiency than low-risk HPV types, and only the high-risk HPV oncoproteins are capable of degrading the p53 proteins.66 In the minority of cervical cancers that are not associated with HPV, p53 is inactivated by mutation. Thus, inactivation of p53, through either E6 binding or mutation, appears to be a central component of the malignant transformation of cells to cervical cancer.62 Mutations in the p53 gene have been identified as the most common genetic abnormality in human tumors.50
The E7 oncoprotein is, like the E6 protein, a small zinc-binding protein. It has approximately 150 amino acids and an amino-terminal domain that is similar in function and sequence to the protein produced by related DNA-transforming viruses (i.e., adenovirus E1A and polyomavirus large tumor antigen). These tumors are all capable of forming complexes with several host cellular proteins, including the retinoblastoma tumor suppressor gene product (pRb).52 The intact retinoblastoma gene is known to protect against tumor development. The congenital form of retinoblastoma is thought to develop when both inherited alleles of the retinoblastoma gene Rb-1 are defective. If one of the defective Rb-1 alleles is inherited, then a somatic point mutation that renders the other one defective will result in the development of retinoblastoma.67 Thus, the intact retinoblastoma gene protects against tumor development. Complex formation of HPV 16 E7 proteins and similar oncogenes inactivates pRb, resulting in deregulated cellular growth.
Besides inactivation and degradation of pRb and related proteins, HPV E7 protein may also interfere directly with key regulatory proteins of the cell cycle, such as cyclin A and cyclin B, which limit transition of the cell from G into S phase. This cellular growth control function seems to be critical in ensuring replication of intact cellular DNA and cell cycle progression. If interfered with, unregulated cell proliferation occurs.68
HPV DNA is detectable in more than 90% of cervical cancers and cases of high-grade dysplasia, and in almost all cases the viral DNA is integrated into the cellular DNA.39, 69 This is in contrast to the benign lesions (i.e., condyloma and most low-grade intraepithelial neoplasms), in which all of the viral DNA is present in the host cell nucleus outside the chromosomes in nonintegrated (episomal) form. During integration, the circular viral DNA is disrupted in a specific region between the E1 and E2 ORFs so that the regulatory function exerted by the E2-derived proteins on the viral genome is lost. This may result in unrepressed DNA transcription of the intact remaining E6 and E7 ORFs.70 The E6 and E7 gene products then mediate proliferative changes in the host cell and cause abnormalities in host cell DNA replication, as described above.
HPV-ASSOCIATED NEOPLASMS
With the advent of molecular probes, HPV DNA has been found in carcinomas and precursor lesions of the anogenital tract, including cervix, vagina, vulva, anus, and penis. HPV DNA has also been found in extragenital Bowen's disease and carcinoma of the urethra, bladder, oral cavity, larynx, esophagus, conjunctiva, cornea, and others. Most of these lesions tested positive for HPV-16.71 Although the data linking HPV to these neoplasms are strong, neither the incidence of these tumors nor the association with HPV approaches that of cervical carcinomas.72
It follows that the uterine cervix is particularly susceptible to HPV infection and malignant transformation. This is because the cervix uniquely meets several requirements for successful transmission of the virus and carcinogenesis. During sexual intercourse, the most commonly HPV-infected part of the penis, the prepuce, is brought into contact with the cervix. Close physical contact during sexual intercourse also places the vagina, vulva, and penis at risk for HPV infection, but evidently not at high risk for neoplasia when compared to the cervix. The heightened vulnerability of the cervix to HPV infection and neoplasia lies in the transformation zone. It is here where more than 80% of cervical neoplasias take their origin, and rarely in the original squamous epithelium. The immature metaplastic epithelium of the transformation zone is mechanically weak and prone to epithelial disruption during sexual intercourse, allowing HPV to attach itself. The infection of the cervix within the transformation zone may be facilitated by specific receptors for HPV called alpha-6 integrins, which are expressed only in basal cells and epidermal stem cells.73
Parkin et al. have attempted to quantitate the role of HPV in human cancer.1 Because oncogenic HPV can be detected by PCR in virtually all cases of cervical cancer, all cases of cervical cancer are "attributed" to HPV infection, which amounts to approximately 492,800 cases per year globally. Using PCR the prevalence of HPV in vaginal cancer is about 60–65% and 20–50% in vulval cancers. There are approximately 40,000 cases of vulval and vaginal cases per year, of which 16,000 are attributable to HPV infection. Ninety per cent of anal cancers are attributable to HPV infection, with a total number of 30,400 cases reported in men and women, of which 27,300 are attributable to HPV infection. About 3% of mouth cancers are attributed to HPV infection, amounting to eight cases per year. Oropharyngeal cancers are attributed HPV infection in about 12% of cases amounting to 6200 cases per year. Overall, the number of cancers attributed to HPV infection globally is 561,100, or about 5.7% of all cancers reported.
EFFECTS OF HPV ON TISSUE
Although squamous epithelium is the principal site of HPV infection, HPV DNA has been isolated from reserve or undifferentiated epithelial cells, which give rise to both the squamous and glandular components of the cervix. This would explain the association of HPV with adenocarcinomas and undifferentiated (small cell) carcinomas of the cervix.74, 75
Latent infection
Infection with HPV is thought to occur when large numbers of virus particles released from infected superficial cells or keratin fragments gain access to basal cells through epithelial breaks in susceptible people. The virus may remain in the basal layer of the epithelium as a separate chromosomal piece of circular DNA termed episome.69 Because the infected cells are histologically and cytologically indistinguishable from uninfected cells, the infection is called latent or occult.
Normal-appearing squamous epithelium adjacent to cervical intraepithelial neoplasia does not commonly contain significant amounts of HPV DNA. It appears that the early occult infection is not maintained outside of epithelial abnormalities. This is consistent with the low recurrence rates that follow ablation as well as the low rate of positive HPV tests in normal cervixes during follow-up.76
The separation between latent and clinically apparent HPV infection is not as precise as previously thought. Low-grade HPV-associated lesions may spontaneously regress, and women with latent HPV infection frequently develop clinically observable lesions over time. Moreover, prospective follow-up studies using sensitive and accurate typing methods have found that the detection of HPV DNA in women without clinically apparent lesions is variable, and that women classified as being latently infected at one visit may be classified as being HPV-negative at a subsequent visit.38
Productive infection
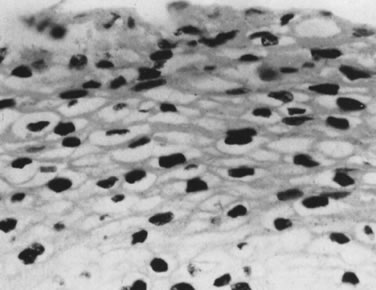
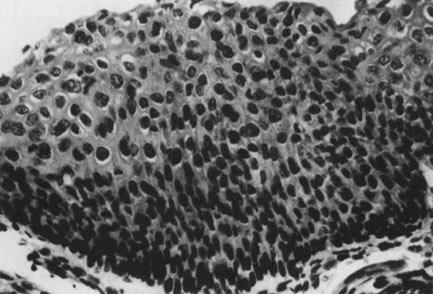
Conditions permitting, viral replication occurs after an incubation period ranging from a few weeks to many years. Replication of episomal DNA is highly restricted in the basal layers and occurs only once per cell cycle. As infected epithelial cells mature and migrate toward the surface, constraints to viral replication are released, and transcription of the late ORF, L1, and L2 occurs, resulting in production of capsid proteins and assembly of infectious virus. This phase of the viral infection is called productive infection and may be associated with a pronounced cytopathic and histopathic effect. The cytopathic effect is most evident in the upper layer of the epithelium and consists of formation of the characteristic koilocytes exhibiting perinuclear cytoplasmic vacuolation, chromatin clumping, and hyperchromasia. Histologic changes may occur and include proliferation of the basal layer (acanthosis), keratin formation (parakeratosis, hyperkeratosis), and capillary overgrowth with formation of papillary projections (papillomatosis), which are pathognomonic for the virus (Fig. 6). On the external genital tract, nonpapillomatous infections are more common than grossly visible papillary changes. The nonpapillomatous epithelial changes are called subclinical infections because they are not recognized by the usual clinical diagnostic methods but instead require exfoliative cytology or magnification for detection.
|
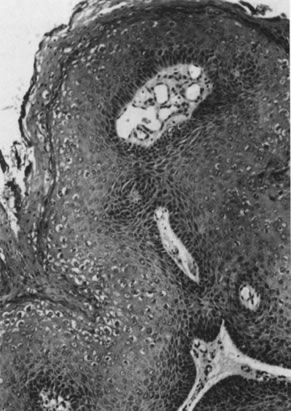
Because of the histologic and etiologic similarity to condylomata acuminata, these flat, koilocytotic changes are commonly termed flat condyloma. The term is a most unfortunate creation because it constitutes a contradiction in terms, meaning “flat protuberance” (from the Greek condyle, meaning knuckle or protuberance). When viewed through the colposcope, the flat condylomas have the same or a similar appearance as dysplasia.
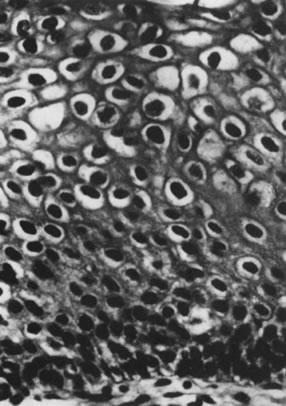
Sometimes, the virus-induced cellular changes are incomplete and focal. These histologic changes have been referred to as borderline koilocytotic atypia and are characterized by perinuclear cytoplasmic clearing, with mild to moderate variation in nuclear size in conjunction with hyperchromaticity and irregularity of the nuclear membranes77 (Fig. 7). Many of these epithelial changes probably represent nonspecific cellular reactions to inflammatory stimulants other than HPV.
|
Nonproductive infection
Under certain conditions, particularly in the presence of potentially oncogenic HPV types such as HPV-16 and HPV-18, the infected cells may fail to differentiate and do not permit completion of the viral cycle. Such nonproductive infections constitute, for the virus, a biologic dead end. They occur in higher-grade dysplasia and invasive cancer. Koilocytotic changes decrease as the degree of dysplasia increases. Basal cell proliferation, nuclear atypia, pleomorphism, atypical mitotic figures, and progressive loss of base-to-surface maturation are recognized as the hallmarks of HPV-associated neoplasia.77 Nonproductive infections of the uterine cervix are, with the exception of invasive cancers, subclinical.
Role of cofactors
The high prevalence of HPV types, including high-risk types in 29% or more of clinically normal women, contrasts with the relatively low incidence of cervical neoplasia.78 Only about 10% of all genital HPV infections come to clinical attention as condyloma or dysplasia, and less than 1% of women with HPV infection develop cervical cancer. Therefore, exposure to HPV alone is not sufficient to produce neoplastic cell transformation. The development of malignant growth appears to be determined by additional factors that modify host cell genes. Several of the following cofactors have been identified in animal and human models.
Malignant conversions of papillomas in rabbits caused by the cottontail rabbit papillomavirus are dependent on treatment with chemical carcinogens (e.g., tar or methylcholanthrene), mechanical scarification, or exposure to ultraviolet radiation. Genetic factors also are likely to play a role because progression to cancer occurs much more frequently and at an earlier age in domestic rabbits than in the natural host, the cottontail rabbit.79, 80 Alimentary tract papillomas in cattle caused by bovine papillomavirus type 4 undergo malignant change only if the cattle graze on pastures with brackenfern. Brackenfern is known to contain radiomimetic substances.81
Patients with epidermodysplasia verruciformis suffer from a defect in cell-mediated immunity so that they cannot render an appropriate immune response to a large number of HPV types (see Table 2). As a consequence, patients with epidermodysplasia verruciformis develop disseminated skin warts resembling reddish plaques, or flat warts in children. Commonly, the warts undergo malignant transformation, particularly in areas exposed to sunlight, indicating a role for ultraviolet irradiation in their development.82
Besides the possible role of smoking and chronic bacterial infection, experimental evidence suggests that viruses such as herpes simplex virus type 1 may mediate changes in HPV-infected cells analogous to chemical and physical carcinogens.83 Other observations have implicated hormones in cervical carcinogens. Indeed, a hormone-responsive element in the noncoding region of genital HPV has been found. Specifically, progesterone stimulation may lead to increased virus production and enhance proliferation of viral DNA-carrying cells. Both events would explain why long-term contraceptive users appear to have a slightly elevated risk of developing cancer of the cervix and why women are more likely to test HPV positive during pregnancy than in their nonpregnant state.83
CLINICAL MANIFESTATIONS
Infection by most sexually transmitted HPV types occurs throughout the lower female genital tract, and multiple sites are commonly involved at the same time. Initially, the total area of infected epithelium greatly exceeds the area displaying lesions. The manifestation of the HPV infection is influenced by cofactors, as previously discussed, and by the viral subtype, involved organ, immune system, and local tissue factors.
Viral subtype
According to their ability to induce cancers, HPV types often are grouped as low-risk types (HPV-6 and -11), medium- or intermediate-risk types (HPV-33, -35, -39, -40, -43, -45, -51, -46, and -58), and high-risk types (HPV-16, -18, and -31). Intermediate- and high-risk viruses also are referred to as potentially oncogenic types. High-risk viruses commonly are isolated from high-grade dysplasias or condylomatous lesions with atypical mitoses. Up to 35% of bland-appearing condylomas, however, may contain potentially oncogenic viruses.79
Involved organ
The HPV-6 and HPV-11 types cause exophytic condylomas (genital warts, condylomata acuminata) mostly on the vulva, vestibule, or perineum, and rarely on the cervix. The same viral types also may be associated with clinically inapparent or subclinical disease, which requires special methods for diagnosis, such as colposcopy. The cervical transformation zone appears particularly vulnerable to oncogenic viruses. Therefore, the most severe effects of high-risk infections generally occur on the cervix.
Immune system
An intact immune system prevents most HPV infections from becoming clinically evident. In immunosuppressed patients, by contrast, HPV virtually always produces recognizable epithelial proliferations, cellular atypia, or neoplasia. Pregnancy is a state of relative immunosuppression, permitting a higher replication rate of viral DNA with enhanced lesion formation and growth.
Local tissue factors
Epithelial abnormalities induced by HPV tend to be localized, although surrounding epithelium also is infected and may exhibit inconspicuous epithelial changes. Localized disease, therefore, may represent a focal breakdown of host control within a field of diffuse HPV infection.52
CERVIX
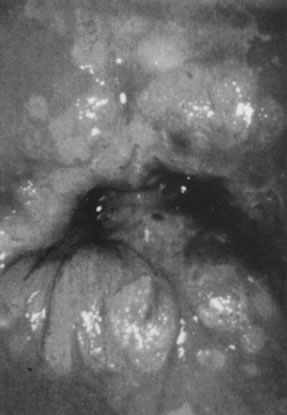
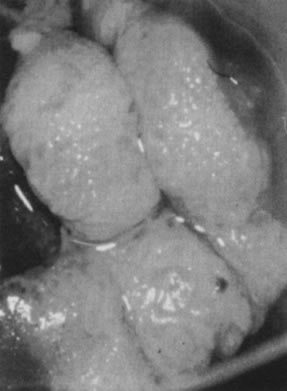
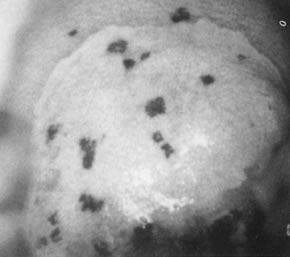
On the cervix, the classic condylomata acuminata associated with HPV-6 and HPV-11 are uncommon and, when present, often are small and best assessed using the colposcope after application of 5% acetic acid. Most HPV-associated lesions are subclinical and present a large variety of colposcopic findings. In some patients, islands of acetowhite epithelium (satellite lesions) are found in the periphery of the cervix outside the transformation zone (Fig. 8).
|
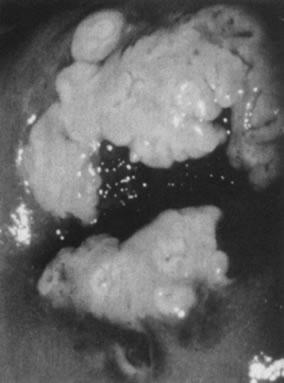
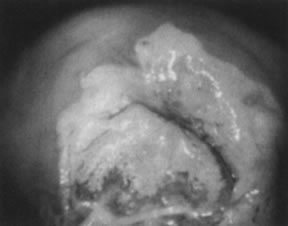
The surface may appear micropapillary (spiked), flat, or sometimes microconvoluted to a brainlike epithelial arrangement (Fig. 9). Many of the flat lesions have a pure, shiny-white color reminiscent of pearls, in contrast to the full, oyster-white color of high-grade cervical intraepithelial neoplasia (Figs. 10 and 11). Some HPV infections produce coarse capillary loops with a horizontal or vertical orientation, giving the appearance of a mosaic or punctation. The regular spacing of the vessels helps to differentiate these arrangements from invasive carcinomas.84, 85 Histologically, the lesions contain koilocytotic and parakeratotic changes in the upper layers of the infected epithelium (Fig. 12).
|
|
|
|

VAGINA
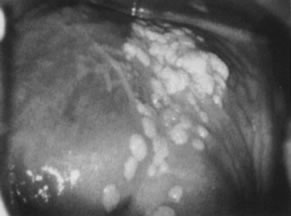
Vaginal condylomata acuminata can be detected in one third of women with vulvar condylomata by careful examination. The distribution usually is patchy, with the proximal and distal thirds of the vagina being affected most commonly. Subclinical vaginal HPV-associated lesions include minute papillary epithelial projections (asperities), each containing a central capillary (Fig. 13). The projections may be isolated, in clusters, or diffuse, covering large areas of the vagina (micropapillomatosis vaginalis). These lesions most commonly are associated with HPV-6 and HPV-11. Acetowhite epithelium, by contrast, is more commonly found in infections due to potentially oncogenic HPV types, most notably HPV-16. Sometimes, minute capillaries are evident, giving rise to patterns reminiscent of punctation and a mosaic pattern on the cervix. Histologically, vaginal intraepithelial neoplasia often is present.86 A common subclinical finding of HPV infection is a pattern called reverse punctation. It consists of a myriad of slightly raised, tiny acetowhite dots, which are pronounced during pregnancy and may involve large areas of the vagina (Fig. 14). Acetic acid may accentuate the changes, but there usually is no sharp demarcation to normal epithelium. It is questionable whether this pattern is due to HPV infection or simply represents a nonspecific response to infectious, hormonal, or other stimuli. If HPV positive, HPV-6 and HPV-11 are isolated from these lesions. Histologic findings are characterized by mild and often focal koilocytosis, variable dyskeratosis, and prominent intraepithelial capillaries.
|
|
VULVA
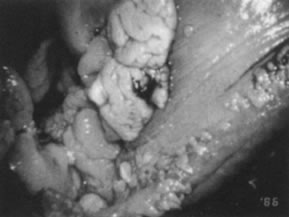
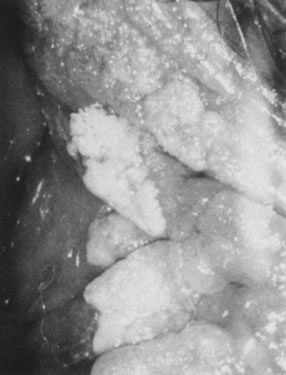
Condylomata acuminata in the vulvar area most commonly involve the perineum, the posterior portion of the vestibule, and the labia minora; less commonly involved are the labia majora, the clitoris, and the mons pubis. Lesions on moist, mucosal surfaces tend to be pink, vascular tumors with finger-like projections (Fig. 15). On keratinized skin, the condylomas are often white or dark because of keratin and pigment formation (Fig. 16). The lesions have a typical histologic appearance of a papillary growth with marked acanthosis, koilocytosis and hyperkeratosis, or parakeratosis. Condylomata acuminata most commonly are associated with HPV-6 and HPV-11.
|
|
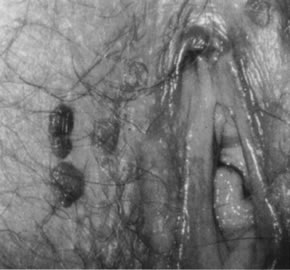
Potentially oncogenic HPV types, especially HPV-16, also may give rise to grossly visible lesions presenting as multiple, darkly pigmented, sometimes white, pale, or fleshy papules, formerly often referred to as bowenoid papulosis (Fig. 17). Histologic examination of the papules usually reveals vulvar intraepithelial neoplasia (Fig. 18).
|
|
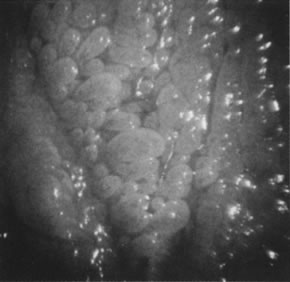
Other HPV-associated lesions of the vulva consist of papillary changes commonly visible with the naked eye but best appreciated colposcopically. The papillae are multiple, small villous projections from mucous membranes that may involve the entire vestibule and inner surface of the labia minora. When extensive, the papillary changes are referred to as micropapillomatosis vestibularis or labialis (Fig. 19). Some women with vestibular papillae have intense vulvar pain (vulvodynia), burning, irritation, and pruritus. Although the papillary formations have been associated with various HPV types, especially HPV-6, they frequently represent anatomic variants of vestibular mucosa.87 It is unlikely that HPV infections are responsible for vulvodynia, but overly aggressive therapy directed against vulvar or vaginal HPV-associated lesions may result in damage to the vulvar epithelium and chronic vulvar pain.
|
Histologically, isolated papillary fronds are observed, with prominent fibrovascular cores associated with chronic inflammation and dilated capillaries. Koilocytotic transformation of superficial epithelial cells is variable.
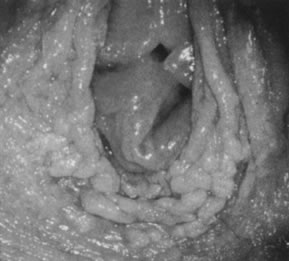
Acetowhitening of the vulvar epithelium occurs most commonly within mucosal surfaces but is not confined to it (Fig. 20). Although it often represents a nonspecific epithelial reaction to chemical, mechanical, and infectious irritants, acetowhite epithelium frequently is associated with HPV-16 and HPV-18, particularly when it is multifocal and when significant epithelial atypia (vulvar intraepithelial neoplasia) is present.
|
NATURAL HISTORY
Progression and spontaneous regression
Women with cytologic evidence of HPV in their Pap smears have a relative risk of developing cervical carcinoma that is 15.6 times higher than the general population.87 Condylomata acuminata can be expected to regress spontaneously in at least 20% of cases. Flat condylomata of the cervix resolve without therapy in 40–50% of cases; 20–40% progress to dysplasia or (rarely) carcinoma, and about 20% persist over an observation period of 1 year.88, 89
Lesions with HPV-6 and HPV-11 are unlikely to develop significant epithelial atypia and are virtually assured a benign course. However, about 30% of flat condylomas and low-grade cervical dysplasias are associated with potentially oncogenic HPV types. Regression of these lesions is uncommon, and progression to invasive carcinoma may occur in up to 5% of cases. The findings emphasize that flat condylomas and low-grade dysplasia have a definite potential for malignant transformation and should not be ignored.90
More than 80% of invasive cervical cancers are preceded by high-grade cervical dysplasia for as long as 20 years. Many women with high-grade dysplasias do not develop cancer, but precise estimates of progression are confounded by length of follow-up and by biopsy altering the natural history of the disease. Estimates of progression of high-grade dysplasia to invasive cancer vary from 20% to more than 70%. The regression rate has been estimated to be as high as 32%.91 Some invasive carcinomas occur in the absence of a previously documented precursor lesion (that is, in the interval between a normal smear and the diagnosis of invasive carcinoma). These malignancies are sometimes referred to as interval cancer. They appear to be more common in younger women (25–35 years of age); are typically small cell carcinomas, adenocarcinomas, and adenosquamous carcinomas; are aggressive in their clinical course; and are strongly related to HPV-18.92 Others have argued that rapidly progressive carcinomas may be a manifestation of endocervical carcinomas that have been inadequately screened.93
Assessment of malignant potential
Histologic examinations of biopsy specimens from infected epithelium provide only a rough guide to the malignant potential of individual lesions. Although atypical mitoses are the hallmark of oncogenic HPV types, atypical mitoses also can be found in some middle-grade lesions caused by nononcogenic HPV types.84
High-risk HPV types commonly cause derangement in chromosomal content, resulting in aneuploidy. Aneuploid lesions rarely regress spontaneously. Polypoid lesions, by contrast, disappear in most cases. Thus, quantitation of cellular DNA content can identify a subset of lesions that are at much greater risk of malignant transformation. A proportion of HPV-16 or HPV-18 infections initially may present as polyploid lesions but subsequently evolve into aneuploid premalignancies.94 In addition, ploidy analyses are cumbersome and not suitable for routine clinical use.
PREVENTION
The high prevalence of latent HPV infections among sexually active men and women will continue to provide an abundant reservoir of HPV-related problems. Given a prevalence of HPV infections of 30% and a transmission rate of 65%, the risk of contracting the virus from sexual contacts with one person may be as high as 20%. Although prospective clinical studies are not available, condoms are likely to provide some protection but are incomplete barriers because they do not shield the scrotum, pubic area, and anus.
Primary prevention of HPV associated disease through vaccination is an exciting new tool in our armamentarium and two prophylactic vaccines have become available globally. HPV infection is cleared by the development of a strong cell-mediated immune response (CMI) that is associated with lesion regression and protection against further infection with the same genotype of HPV. Humoral (antibody) immunity is evoked in most, but not all infected individuals and is directed against the major capsid protein L1 displayed on the outer surface of the intact virus particle. This antibody is considered to be virus neutralizing, but serum-neutralizing antibodies to natural infection are low, probably due to the exclusively intraepithelial nature of HPV infection, with the absence of a viremia. With this in mind, it was thought that vaccines that could generate neutralizing antibodies to HPV capsid antigen might be effective in prophylactically. L1 proteins can be expressed via recombinant yeast or viral vectors and the L1 protein can self assemble into empty capsids or virus-like particles (VLPs) that are morphologically and antigenically identical to native virions. This technology has been used to develop two commercially available HPV vaccines. Cervarix is a bivalent HPV-16/18 VLP vaccine developed by GlaxoSmithKline. In this vaccine the L1 protein of each HPV type is expressed by a recombinant baculovirus vector. The L1 VLPs are formulated with a ASO4 adjuvant comprising 500 μg of aluminium hydroxide and 50 μg of 3-deacylated monophosphoryl lipid A. Gardasil is a quadrivalent HPV-16/18/6/11 L1 VLP vaccine developed by Merck and Co, Inc. The L1 protein for each HPV VLP type is expressed via a recombinant Saccharomyces pombe vector. Both vaccines are administered by three intramuscular injections over a 6 month period.95
Results from clinical trials (phase 2 and 3) have been reported.96, 97, 98 All trials were blinded, randomized, and placebo controlled of young women (mean age 20 years). The prophylactic efficacies of the vaccines against the HPV types targeted have been shown to greater than 95% against all reported vaccine-type specific endpoints, which include incident HPV infection, persistent infection, and preinvasive squamous lesions of the anogenital tract and genital warts in the case of the quadrivalent vaccine. Both vaccines showed an excellent safety and tolerability profile.
Ideally the vaccine should be administered to individuals prior to the onset of sexual activity, as the vaccine is prophylactic and not therapeutic. Ideal age of vaccination will differ in different communities but in most the target age group will be girls (and boys depending on resources) aged 9–15 years of age, with "catch up" vaccination of individuals aged 16–26 years. In the coming years, more information on the performance of the vaccine in males, mature women, immunosuppressed populations, and young women with prior HPV exposure will become available. The key unanswered question in young women is whether the initial vaccination will provide lifelong protection from disease associated with the targeted HPV types or whether booster doses will be required.
PRINCIPLES OF MANAGEMENT
- Establish the diagnosis. Biopsy is required in all cases, except for the most obvious classic condylomata acuminata.
- Assess the extent of disease. Colposcopy and acetic acid application should be part of the initial assessment of HPV-associated epithelial changes, particularly when tissue destruction and ablation techniques, such as cryotherapy, local excision, electrocautery, or laser, are used. Colposcopy and other forms of examination under magnification are also useful during follow-up.
- Treat or eliminate associated problems. Immunosuppression related to wasting disease, diabetes mellitus, diseases of the immune system, stress, drugs, and hormones may strongly affect the course of HPV infections. Although avoidance of stress, better control of diabetes mellitus, and improvement of the nutritional status of patients with wasting disease often are feasible and effective, immunosuppressive drugs rarely can be discontinued. Most gynecologists also are reluctant to discontinue oral contraceptives because the danger associated with pregnancy usually outweighs the potential benefits of reduced HPV DNA replication. Treating coexistent trichomoniasis may also be helpful.
- Examine and treat the partner. Successful treatment of the male sexual partner of an infected woman may prevent future sex partners from becoming infected. There is no good evidence to suggest that male partner examination and treatment alters the course of the disease in the infected female partner.99
- Do no harm. Not all patients with HPV infections require treatment. The physician must consider the possibility of spontaneous regression and weigh the discomfort and risks of therapy against the existing disability.
- Select proper treatment. In choosing treatment for a particular patient, the physician should consider the clinical presentation and manifestations of the HPV infection and the effectiveness, availability, ease of application, side effects, long-term risks, and cost of the treatment method. The patient's tolerance and preferences, suitability for use in pregnancy, vaginal application, and control of latent infection also are of concern.
INDICATIONS FOR TREATMENT
Major indications
ALLEVIATE IRRITATION, DISCOMFORT, OR PAIN
Patients with symptomatic condylomata acuminata fall into this category.
PREVENT NEOPLASIA
The known association of HPV with squamous neoplasia of the lower genital tract requires that precursor lesions be eradicated. Although most genital dysplasias do not transform into invasive carcinoma, there is general agreement to treat patients with high-grade (moderate and severe) dysplasia and carcinoma in situ. The role of therapy for low-grade lesions (flat condyloma, mild dysplasia) is less well defined. In view of the high rate of spontaneous regression of minor HPV-associated epithelial abnormalities, a watch-and-wait approach appears appropriate. For cervical lesions, the inconvenience, anxiety, time, and cost involved in repeated cytologic testing, colposcopy, and biopsies should be weighed against the relatively innocuous, effective ablation of the transformation zone by cryotherapy or other methods in the office.
Minor indications
CONTROL SPREAD OF DISEASE
Although most genital HPV-associated lesions are not genuine precursors of carcinoma, effective treatment may help to bring the troublesome HPV epidemic under control. Patients with asymptomatic HPV infections may be treated for this reason. Ablation of the products of HPV infection, such as exophytic or flat condylomata, however, by no means eliminates the virus. No method has been developed to eradicate latent or subclinical HPV infections, which, as previously emphasized, tend to involve the entire lower genital tract.100
ALLEVIATE ANXIETY
Ablation of an abnormal transformation zone by cryotherapy reduces the risk of cervical carcinoma and may alleviate anxiety. The patient should be advised, however, that the virus is likely to persist in the periphery of the cervix and elsewhere in the lower genital tract, that she will remain potentially infectious, and that the virus probably cannot be completely eradicated. The understanding that HPV infections are extremely common and that the cancer risk in other areas of the genital tract and in male partners is very small should further alleviate mental anguish. The need for continued screening even after ablation of the transformation zone should be emphasized.
TREATMENT
The ideal therapy for HPV-associated lesions should be effective, readily available, easily applied, well tolerated, free of long-term risks, safe in pregnancy, suitable for vaginal use, and inexpensive. It should also take into consideration that genital HPV infection is a disease of both normal- and abnormal-appearing epithelium that often runs its course over several weeks to 8 months and spontaneously disappears. Therefore, the surgical approach to therapy is less than optimal. Destruction of the products of HPV infections, such as condylomas or dysplasias, leads to remission in about 85% of patients despite of the fact that HPV-associated lesions are often surrounded by areas of subclinical infection. Consequently, treatment methods with the potential to control subclinical disease, such as laser, fluorouracil (5-FU), and interferon, should be reserved for patients whose lesions are too extensive for control by caustic agents or outpatient destruction and for those who do not respond to simpler therapies.81
The carbon dioxide laser is considered the most effective form of treatment of extensive vulvar HPV-associated lesions. A trial with periodic 5-FU may still be warranted, however, because about 70% of patients with extensive HPV-associated disease respond to this regimen and may be spared the expense of laser therapy, which usually requires general anesthesia.101 The control rate of subclinical disease and the overall cure rate appear to be higher when laser is used in association with adjuvant (prophylactic) 5-FU.102 5-FU is probably superior to laser for the treatment of vaginal condylomata acuminata and is at least as effective as laser for multifocal vaginal dysplasia.103, 104, 105
In the future, antiviral drugs or immune modulators may be more desirable, and interferons were the first such agents to show promise. The efficacy of systematic interferon demonstrated so far may not warrant the toxic reactions observed, however, and the efficacy of topical interferon remains in doubt. Intralesional injection is no more convenient than destructive modalities and, unless subclinical virus is proved to be eliminated, shows no obvious therapeutic advantage.105
Imiquimod, an immune-response modifier supplied in a cream base, is applied by patients with condylomata acuminata. It is the first successful treatment that promotes a host immune response, resulting in complete clearance of the lesions in more than 50% of women treated.
Highly specific treatments for HPV-associated lesions would be based on the known biologic properties of HPVs and may consist of targeting oncoproteins expressed in malignant HPV-associated lesions. Experiments with antisense oligonucleotides targeting the E6 and E7 oncoproteins showed growth inhibition of cancer cell lines.106, 107 At present, problems with cellular transfer and targeting using recombinant DNA technologies severely limit their clinical applications.
ACKNOWLEDGMENT
I thank Sonia M. Kheir for contributing to this chapter.
REFERENCES
Parkin DM, Bray F: The Burden of HPV-related cancers. Vaccine 24(Suppl 3): S11–25, 2006 |
|
Rous P, Beard JW: The progression of carcinoma of virus-induced rabbit papillomas (Shope). J Exp Med 62: 523, 1935 |
|
Meisels A, Fortin R: Condylomatous lesions of cervix and vagina. I. Cytologic patterns. Acta Cytol 20: 505, 1976 |
|
Shope RE: Infectious papillomatosis of rabbits. J Exp Med 58: 607, 1933 |
|
Purola E, Savia E: Cytology of gynecologic condyloma acuminatum. Acta Cytol 21: 26, 1977 |
|
Koss LG, Durfee GR: Unusual patterns of squamous epithelium of the uterine cervix: Cytologic and pathologic study of koilocytotic atypia. Ann NY Acad Sci 63: 1245, 1956 |
|
Della Torre G, Pilotti S, DePalo G et al: Viral particles in cervical condylomatous lesions. Tumori 64: 459, 1978 |
|
Laverty CR, Booth N, Hills E et al: Noncondylomatous wart virus infection of the postmenopausal cervix. Pathology 10: 373, 1978 |
|
Jenson AB, Rosenthal JD, Olson C et al: Immunological relatedness of papillomavirus from different species. JNCI 64: 495, 1980 |
|
Shah KV, Lewis MG, Jenson AB et al: Papillomavirus and cervical dysplasia. Lancet 2: 1190, 1980 |
|
Wright TC, Richart R: Role of human papillomavirus in the pathogenesis of genital-tract warts and cancer. Gynecol Oncol 37: 151, 1990 |
|
Trottier H, Franco EL: The epidemiology of genital human papillomavirus infection. Vaccine 24(Suppl 1): S4–S15, 2006 |
|
de Sanjose S, Diaz M, Castellsague X et al: Worldwide prevalence and genotype distribution of cervical human papillomavirus DNA in women with normal cytology; a meta-analysis. Lancet Infect Dis 7: 453–59, 2007 |
|
Kjaer SK, Svare EI, Worm AM et al: Human papillomavirus infection in Danish female sex workers. Decreasing prevalence with age despite continuously high sexual activity. Sex Transm Dis 27: 438–445, 2000 |
|
Bauer H, Ting Y, Greer CE et al: Genital human papillomavirus infection in female university students as determined by a PCR based method. JAMA 265: 472, 1991 |
|
Koutsky LA, Holmes KK, Critchlow CW et al: A cohort study of the risk of cervical intraepithelial neoplasia. grade 2 or 3 in relation to papillomavirus infection. N Engl J Med 327: 1272, 1992 |
|
Sanjose S, Diaz M, Castellsague X et al: Worldwide prevalence and genotype distribution of cervical human papillomavirus DNA in women with normal cytology: a meta-analysis. Lancet 7: 453, 2007 |
|
Fairley CK, Chen S, Ugoni A et al: Human papillomavirus infection and its relationship to recent and distant sexual partners. Obstet Gynecol 84: 755, 1994 |
|
Ho GY, Bierman R, Beardsley L, Chang CJ, Burk RD: Natural history of cervicovaginal papillomavirus infection in young women. N Engl J Med 338: 423–8, 1998 |
|
Winder RL, Lee SK, Hughes JP, Adam DE, Kiviat NB, Koutsky LA. Genital human papillomavirus infection: incidence and risk factors in a cohort of female university students. Am J Epidemiol 157: 218–26, 2003 |
|
Woodman CB, Collins S, Winter H et al: Natural history of cervical human papillomavirus infection in young women: a longitudinal cohort study. Lancet 357: 1831–6, 2001 |
|
Franco ElL, Villa LL, Sobrinho JP et al: Epidemiology of acquisition and clearance of cervical human papillomavirus infection in women from a high-risk area for cervical cancer. J Infect Dis 180: 1415–23, 1999 |
|
Mosciki AB, Shiboski S, Broering J et al: The natural history of human papillomavirus infection as measured by repeated DNA testing in adolescent and young women. J Pediatr 132: 277–84, 1998 |
|
Molano M, van den Brule A, Plummer M et al: Determinants of clearance of human papillomavirus infections in Colombian women with normal cytology: a population-based, 5 year follow-up study. Am J Epidemiol 158: 486–94, 2003 |
|
Liaw KL, Glass AG, Manos MM et al. Detection of human papillomavirus DNA in cytologically normal women and subsequent cervical squamous intraepithelial lesions. J Natl Cancer Inst 91: 954–60, 1999 |
|
Edwards S, Carne C: Oral sex and the transmission of viral STIs. Sex Transm Infect 74: 6–10, 1998 |
|
Sonnex C, Strauss S, Gray JJ. Detection of human papillomavirus DNA on the fingers of patients with genital warts. Sex Transm Infect 75: 317–319, 1999 |
|
Palesfsky JM, Holly EA, Ralston ML et al: High incidence of anal high-grade squamous intra-epithelial lesions among HIV-positive and HIV negative homosexual and bisexual men. AIDS 12: 495–503, 1998 |
|
Wickenden C, Hanna N, Taylor Robinson D et al: Sexual transmission of human papillomaviruses in heterosexual and male homosexual couples, studied by DNA hybridization. Genitourin Med 64: 34, 1988 |
|
Krebs HB, Schneider V: Human papillomavirus associated lesions of the penis: Colposcopy, cytology and histology. Obstet Gynecol 70: 299, 1987 |
|
Pakarian F, Kaye J, Cason J et al: Cancer-associated papillomaviruses: Perinatal transmission and persistence. Br J Obstet Gynaecol 101: 514, 1994 |
|
Winer RL, Koutsky LA: Delivering reassurance to parents: Perinatal human papillomavirus transmission is rare. Sex Transm Dis 31: 63–64, 2004 |
|
Walboomers JM, Jacobs MV, Manos MM et al. Human papillomavirus is a necessary cause of invasive cervical cancer worldwide. J Pathol 189: 12, 1999 |
|
Winer RL, Lee SK, Hughes JP et al. Genital human papillomavirus infection: incidence and risk factors in a cohort of female university students. Am J Epidemiol 157: 218–26, 2003 |
|
Smith JS, Herrero R, Bosetti C, Munoz N et al. Herpes simplex virus-2 as a human papillomavirus /cofactor in the etiology of invasive cervical cancer. J Natl Cancer Inst 94: 1604–13, 2002 |
|
Castellsague X, Munoz N: Chapter 3: Cofactors in factors in human papillomavirus carcinogenesis - role of parity, oral contraceptives and tobacco smoking. J Natl Cancer Inst Monogr 31: 20, 2003 |
|
Coggin JR, zur Hausen H: Workshop on the papillomaviruses and cancer. Cancer Res 39: 545, 1979 |
|
Wright TC: Pathogenesis and diagnosis of preinvasive lesions of the lower genital tract. In Hoskins WJ, Perez C, Young RC (eds): Principles and Practice of Gynecologic Oncology, pp 744–752. 3d ed. Philadelphia, Lippincott William & Wilkins, 2000 |
|
zur Hausen, Schneider A: The role of papillomaviruses in human anogenital cancer. In Salzman HP, Howley PM (eds): The Papovaviridiae, p 245. New York, Plenum Press, 1987 |
|
Gissmann L, zur Hausen H: Partial characterization of viral DNA from human genital warts (condylomata acuminata). Int J Cancer 25: 605, 1980 |
|
Munoz, NF. Bosch X, Sanjose S et al: Epidemiologic Classification of Human Papillomavirus Types Associated with Cervical Cancer. N Engl J Med 348: 518–27, 2003 |
|
Bonnez W: Murine models of human papillomavirus-infected human xenografts. Papillomavirus Rep 9: 27, 1998 |
|
Meyers C, Frattini MG, Hudson JB et al: Biosynthesis of human papillomavirus from a continuous cell line upon epithelial differentiation. Science 257: 971, 1992 |
|
Galloway DA: Papillomavirus capsids: a new approach to identify serologic markers of HPV infection. J Natl Cancer Inst 86: 474, 1994 |
|
Rose RA, Bonnez W, Reichman RC et al: Expression of human papillomavirus type 11 L1 protein in insect cells: in vivo and in vitro assembly of virus like particles. J Virol 67: 1936, 1993 |
|
Dürst M, Gissman L, Ikenberg H et al: A papillomavirus DNA from a cervical carcinoma and its prevalence in cancer biopsy samples from different geographic regions. Proc Natl Acad Sci USA 80: 3812, 1983 |
|
Boshart M, Gissman L, Ikenberg H et al: A new type of papillomavirus DNA, its presence in genital cancer and in cell lines derived from genital cancer. EMBO J 3: 1151, 1984 |
|
Castle PE, Lorincz AT, Mielzynska-Lohnas I et al: Results of human papillomavirus DNA testing with the hybrid capture 2 assay are reproducible. J Clin Microbiol 40: 1088–1090, 2002 |
|
Iftner T, Villa LL: Chapter 12: Human Papillomavirus Technologies J Natl Cancer Inst Monographs No. 31: 80–87, 2003 |
|
Vogelstein B, Kinzler KE: p53 function and dysfunction. Cell 70: 523, 1992 |
|
zur Hausen H: Papillomavirus infections: A major cause of human cancers. Biochem Biophys Acta 1288: F55, 1996 |
|
Münger K, Werness BA, Dyson N et al: Complex formation of human papillomavirus E7 proteins with the retinoblastoma tumor suppressor gene product. EMBO J 8: 4099, 1989 |
|
Melkert PWJ, Hopman E, van Den Brule AJC et al: Prevalence of HPV in cytomorphologically normal cervical smears as determined by polymerase chain reaction is age-dependent. Int J Cancer 53: 919, 1993 |
|
Kjaer SK, Engholm G, Teisen C et al: Risks factor for cervical human papillomavirus and herpes simplex virus infection in Greenland and Denmark: A population-based study. Am J Epidemiol 131: 669, 1990 |
|
Arbyn M, Sasieni P, Meijer CJ et al: Chapter 9: Clinical applications of HPV testing: A summary of meta-analyses. Vaccine 24(Suppl3): S78–89, 2006 |
|
Arbyn M, Buntinx F, Ranst M et al: Virologic versus cytologic triage of women with equivocal Pap smears; a meta-analysis of the accuracy to detect high-grade intraepithelial noeplasia. J Natl Cancer Inst 96: 280–93, 2004 |
|
Arbyn M, Paraskevaidis E, Martin-Hirsch P et al: Clinical utility of HPV DNA detection: triage of minor cervical lesions, follow up of women treated for high-grade CIN: an update of pooled evidence. Gynecol Oncol 99(3 Suppl 1): S7–11, 2005 |
|
Franco EL: Primary screening of cervical cancer with human papillomavirus test. J Natl Cancer Inst Monogr 31: 89–96, 2003 |
|
The Atypical Squamous Cells of Undetermined Significance/Low-Grade Squamous Intraepithelial Lesions Triage Study (ALTS) Group. Human papillomavirus testing for triage of women with cytologic evidene of low-grade squamous intra-epithelial lesions: baseline data from a randomised trial. J Natl Cancer Inst 92: 397–402, 2000 |
|
Cox JT, Lörincz AT, Schiffman MH et al: Human papillomavirus testing by hybrid capture appears to be useful in triaging women with a cytologic diagnosis of atypical squamous cells of undetermined significance. Am J Obstet Gynecol 72: 946, 1995 |
|
Howley PM: Role of human papillomaviruses in human cancer. Cancer Res 51: 5019, 1991 |
|
Park DJ, Wilczynski SP, Paquette RL et al: Mutations in HPV-negative cervical carcinoma. Oncogene 9: 205, 1994 |
|
Gissman L, Dürst M, Oltersdorf T et al: Human papillomaviruses and cervical cancer. In Steinberg BM, Brandsma TL, Tacchman LB (eds): Cancer Cells, vol V, The Papilloma Viruses, p 275. New York, Cold Spring Harbor Laboratory, 1987 |
|
Pecoraro G, Morgan D, Defendi D: Differential effects of human papillomaviruses 6, 16, and 18 DNA's on immortalization and transformation of human cervical cells. Proc Nat Acad Sci USA 86: 563, 1989 |
|
Levine AJ: p53, cellular gatekeeper for growth and division. Cell 88: 323, 1997 |
|
Howley PM: Papillomavirinae: The viruses and their replication. In Fields BN, Knipe DM, Howley PM (eds): Fundamental Virology, p 947. Philadelphia, PA, Lippincott-Raven, 1966 |
|
Horowitz JM, Yandall DW, Park SH et al: Point mutational inactivation of the retinoblastoma antioncogene. Science 243: 937, 1989 |
|
Park TW, Fujawara H, Wright TC: Molecular biology of cervical cancer and its precursors. Cancer 76: 1902, 1995 |
|
Dürst M, Kleinheinz A, Holz M et al: The physical state of human papillomavirus type 16 DNA in benign and malignant genital tumors. J Gen Virol 66: 1515, 1985 |
|
Smotkin D: Virology of human papillomavirus. Clin Obstet Gynecol 32: 117, 1989 |
|
Clavel C, Huu VP, Durlach AP et al: Mucosal oncogenic human papillomaviruses and extragenital Bowen's disease. Cancer 86: 282, 1999 |
|
Alani RM, Munger K: Human papillomaviruses and associated malignancies. J Clin Oncol 16: 330, 1998 |
|
Evander M, Frazer IH, Payne E et al: Identification of the alpha-6 integrin as a candidate receptor for papillomaviruses. J Virol 71: 2449, 1997 |
|
Stoler MH, Mills SE, Gersell DJ, Walker AN: Small cell neuroendocrine carcinoma of the cervix: A human papillomavirus type 18 associated cancer. Am J Pathol 15: 28, 1991 |
|
Smotkin D, Berek JS, Fu YS et al: Human papillomavirus deoxyribonucleic acid in adenocarcinoma and adenosquamous carcinoma of the uterine cervix. Obstet Gynecol 68: 241, 1986 |
|
Tate JE, Resnick M, Sheets E, Crum CP: Absence of papillomavirus DNA on normal tissue adjacent to most cervical intraepithelial neoplasms. Obstet Gynecol 88: 257, 1996 |
|
Nuovo GH, Nuovo MA, Cottral S et al: Histological correlates of clinically occult human papillomavirus infection of the cervix uteri. Am J Surg Pathol 12: 198, 1988 |
|
Schneider A, Holz M, Gissmarm L: Prevalence of genital HPV infections in pregnant women. Int J Cancer 40: 198, 1987 |
|
Campion MJ, Franklin EW, Stacy LD et al: Human papillomavirus and anogenital neoplasia: A fresh look at the association. South Med J 82: 35, 1989 |
|
Fischer RG, Silverton J: The virus-induced papilloma-to-carcinoma sequence. IV. Carcinoma in domestic rabbits infected while in uterus. Cancer Res 11: 737, 1951 |
|
Jarrett WFH: The natural history of bovine papillomavirus infections. Adv Virol Oncol 5: 83, 1985 |
|
Orth G, Jablonska S, Favre M et al: Epidermodysplasia verruciformis: A model for viral oncogenesis in man. Cold Spring Harbor Conf Cell Prolif 7: 259, 1980 |
|
zur Hausen H: Papillomaviruses as carcinomaviruses. Adv Viral Oncol 8: 1, 1989 |
|
Reid R, Greenberg M, Jenson AB et al: Sexually transmitted papillomaviral infections. I. The anatomic distribution and pathologic grade and neoplastic lesions associated with different viral types. Am J Obstet Gynecol 156: 212, 1987 |
|
Reid R, Campion MJ: HPV-associated lesions of the cervix: Biology and colposcopic features. Clin Obstet Gynecol 32: 157, 1989 |
|
Campion MJ: Clinical manifestations and natural history of genital human papillomavirus infections. Obstet Gynecol Clin North Am 14: 363, 1987 |
|
Bergeron C, Ferenczy A, Richart RM et al: Micropapillomatosis labialis appears unrelated to human papillomavirus. Obstet Gynecol 70: 281, 1990 |
|
Mitchell H, Drake M, Medley G: Prospective evaluation of risk of cervical cancer after cytological evidence of human papillomavirus infection. Lancet 2: 573, 1986 |
|
Nash JD, Burke TW, Hoskins WJ: Biologic course of cervical human papillomavirus infection. Obstet Gynecol 69: 160, 1987 |
|
Evans AS, Monaghan JM: Spontaneous resolution of warty atypia: The relevance of clinical and nuclear DNA features. A prospective study. Br J Obstet Gynaecol 92: 165, 1985 |
|
Campion MJ, McCance DJ, Cuzick J et al: Progressive potential of mild cervical atypia: Prospective cytological, colposcopic, and virologic study. Lancet 2: 168, 1986 |
|
Ostör AG: Natural history of cervical intraepithelial neoplasia: A critical review. Int J Gynecol Pathol 12: 186, 1993 |
|
Crum CP, Cibas ES, Lee KR: Pathology of Early Cervical Neoplasia, Chap 2, p 7. New York, Churchill Livingstone, 1997 |
|
Schwartz PE, Hadjimichael O, Lowell DM et al: Rapidly progressive cervical cancer: The Connecticut experience. Am J Obstet Gynecol 175: 1105, 1996 |
|
Stanley M, Lowy DR, Frazer I: Chapter 12: Prophylactic HPV vaccines: Underlying mechanisms. Vaccine 24(Suppl 3): 106–113, 2006 |
|
The FUTURE II Study Group: Quadrivalent vaccine against human papillomavirus to prevent high-grade cervical lesions. N Engl J Med 356: 1915–27, 2007 |
|
Harper DM, Fracno EL, Wheeler CM et al: Sustained efficacy up to 4.5 years of a bivalent L1 virus-like particle vaccine against human papillomavirus types 16 & 18 in young women: follow up from a randomised control trial. Lancet 367: 1247–55, 2006 |
|
Ault KA: Effect of prophylactic human papillomavirus L1 virus-like particle vaccine on risk of cervical intraepithelial neoplasia grade 2, grade 3 and adenocarcinoma in situ: a combined analysis of four randomised clinical trials. Lancet 369: 1861–8, 2007 |
|
Lancaster WD, Jenson AB: Human papillomavirus infection and anogenital neoplasia: Speculations for the future. Obstet Gynecol Clin North Am 14: 601, 1987 |
|
Krebs HB, Helmkamp BF: Does the treatment of genital condylomata acuminata in men decrease the treatment failure rate of cervical dysplasia in the female sexual partner? Obstet Gynecol 76: 660, 1990 |
|
Riva JM, Sedlacek TV, Cunnane MF, Mangan CE: Extended carbon dioxide laser vaporization in the treatment of subclinical papillomavirus infection of the lower genital tract. Obstet Gynecol 73: 25, 1989 |
|
Krebs HB: Treatment of extensive vulvar condylomata acuminata with topical 5-fluorouracil. South Med J 83: 762, 1990 |
|
Krebs HB: Combination of laser plus 5-fluorouracil for the treatment of extensive genital condylomata acuminata by weekly topical application of 5-fluorouracil. Obstet Gynecol 70: 68, 1987 |
|
Krebs HB: Treatment of vaginal condylomata acuminata by weekly topical application of 5-fluorouracil. Obstet Gynecol 70: 68, 1987 |
|
Krebs HB: Treatment of vaginal intraepithelial neoplasia with laser and topical 5-fluorouracil. Obstet Gynecol 73: 657, 1989 |
|
Ferenczy A: Comparison of 5-fluorouracil and CO2 laser for treatment of vaginal condylomata. Obstet Gynecol 64: 773, 1984 |
|
Kirby P. Editorial. JAMA 259: 570, 1988 |