This chapter should be cited as follows:
Aurélio Palazzi Sáfadi M, Glob Libr Women's Med
ISSN: 1756-2228; DOI 10.3843/GLOWM.419493
The Continuous Textbook of Women’s Medicine Series – Obstetrics Module
Volume 17
Maternal immunization
Volume Editors:
Professor Asma Khalil, The Royal College of Obstetricians and Gynaecologists, London, UK; Fetal Medicine Unit, Department of Obstetrics and Gynaecology, St George’s University Hospitals NHS Foundation Trust, London, UK
Professor Flor M Munoz, Baylor College of Medicine, TX, USA
Professor Ajoke Sobanjo-ter Meulen, University of Washington, Seattle, WA, USA

Chapter
Zika Virus
First published: May 2023
Study Assessment Option
By answering four multiple-choice questions (randomly selected) after studying this chapter, readers can qualify for Continuing Professional Development points plus a Study Completion Certificate from GLOWM.
See end of chapter for details.

INTRODUCTION
Zika virus (ZIKV) is a single-stranded, positive-sense enveloped RNA arthropod-borne virus (arbovirus) whose genome has one open reading frame. ZIKV belongs to the Flavivirus genus and the Flaviviridae family, which also include other important pathogens such as dengue (DENV), Japanese encephalitis (JE), tick-borne encephalitis (TBE), West Nile (WN), and yellow fever (YF) viruses. Through phylogenetic analysis ZIKV has been characterized into two major lineages namely the African and Asian lineages. ZIKV African lineage has been isolated sporadically in non-human and human specimens since 1947. Asian lineage viruses were first isolated in Malaysia in 1951 and later in the Pacific Islands from 2007 onwards, with some activity in other Western Pacific and Southeast Asia countries since that time. The 2015–16 epidemic in the Americas was caused by a strain of the Asian lineage commonly referred to as the contemporary American strain.1,2
The mature ZIKV particles consist of 90 E homodimers and 90 M homodimers on the lipid membrane, and the genomic RNA is surrounded by C protein (Figure 1). E proteins are responsible for receptor binding, attachment, viral entry, and membrane fusion.3 The ZIKV E protein is composed of three ectodomains (DI, DII, and DIII) and is the primary target of neutralizing antibodies (nAbs).
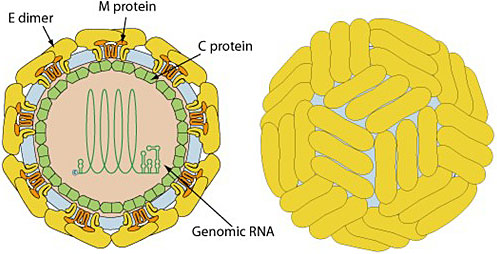
1
Schematic diagram of ZIKV structure. Zika virions are enveloped, spherical, and approximately 50 nm in diameter. The surface M and E proteins are symmetrically arranged on the viral surface. M, membrane; E; envelope; C, capsid. Pictures from ViralZone, SIB Swiss Institute of Bioinformatics (http://viralzone.expasy.org/6756?outline=all_by_protein).
ZIKV genome-encoded polyprotein can be cleaved into three structural proteins – capsid (C), anterior membrane (prM), and envelope (E) – and seven non-structural proteins (NS1, NS2A, NS2B, NS3, NS4A, NS4B, and NS5)3 (Figure 2).
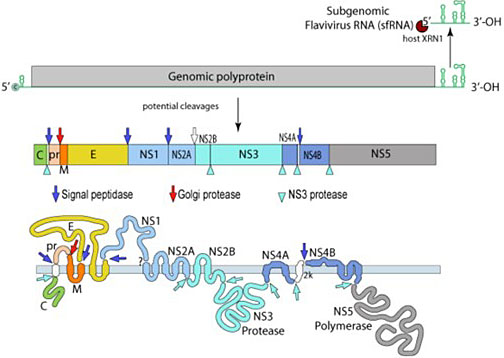
2
The ZIKV genome. RNA containing a single open reading frame that encodes three structural and seven nonstructural proteins. Pictures from ViralZone, SIB Swiss Institute of Bioinformatics (http://viralzone.expasy.org/6756?outline=all_by_protein).
The ZIKV was first isolated in 1947, from a rhesus monkey, in the Zika Forest, Uganda, in a specimen that was being investigated with the purpose of identifying yellow fever virus activity in that region.4 For approximately 60 years after being described, the ZIKV was only reported in sporadic cases in Africa and Asia.5
However, since 2007, a series of virus outbreaks have been reported outside of Africa and Asia: first, in the Federal States of Micronesia, and, subsequently, in French Polynesia and other Pacific Islands.5 In May 2015, autochthonous transmission of ZIKV, associated with an outbreak of “dengue-like syndrome” cases, was detected in the North-Eastern region of Brazil. The ZIKV outbreak continued to evolve, spreading geographically very rapidly.6 Since then, transmission has been identified in almost all countries in the Americas, except mainland Chile, Uruguay, and Canada.
Cases of Zika virus disease globally declined from 2017 onwards; however, Zika virus transmission persists at low levels in several countries in the Americas, remaining the WHO Region with the highest number of reported ZIKV disease cases annually, as well as in other endemic regions. In addition, the first local mosquito-transmitted Zika virus disease cases were reported in Europe in 2019 (France) and Zika virus outbreak activity was detected in India in 2021. By March 2022, a total of 89 countries and territories had reported evidence of mosquito transmitted Zika virus infection; however, surveillance remains limited globally. Worldwide, 61 countries and territories have evidence of established competent Aedes aegypti vectors but so far have not documented ZIKV transmission, anticipating the theoretical potential risk for the occurrence of ZIKV outbreaks in these places.6 (Figure 3).
During the 2015–2016 outbreak, Brazil was the most affected country in the world, with a peak of 216,207 reported cases in 2016. After the exploding rise in the number of cases between 2015 and 2016, there was a significant decrease to only approximately 10,000 cases in 2019, 7000 in 20207, 6000 in 2021 and 9200 in 2022.7 Between 2015 and 2022, the Brazilian Ministry of Health confirmed 1857 cases of congenital ZIKV syndrome, most of them (>80%) diagnosed between 2015 and 2016, predominantly in the North-Eastern region of the country.8
One intriguing aspect of the ZIKV epidemic was the consistent decline in case numbers since 2017, as reported in most affected countries in the Americas. The most likely hypothesis for this phenomenon is that the population would have developed a “herd immunity”, which reduced the number of susceptible, naive subjects, which in turn reduced transmission during seasonal activity.6 However, it is likely that reemergence of Zika virus outbreaks will occur once population immunity wanes and the proportion of naïve individuals increases. According to reports from the PAHO, in 2021, only Brazil, Mexico, Bolivia and Puerto Rico reported confirmed cases of Zika in the Americas region.9
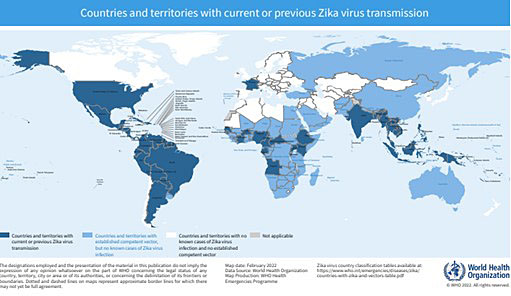
3
World map of Countries and territories with current or previous Zika virus transmission. Picture from WHO – Zika Travel Information (https://cdn.who.int/media/docs/default-source/documents/emergencies/zika/map-of-countries_with_zika_transmission_feb2022.pdf?sfvrsn=802a352a_5).
TRANSMISSION
ZIKV is transmitted to humans primarily by Aedes aegypti mosquitoes (and less commonly by other Aedes species, such as Aedes polynesiensis, Aedes hensilli, Aedes africanus, and Aedes albopictus), the same vector that can transmit dengue, chikungunya, and yellow fever viruses.10 ZIKV has already been isolated from other non-Aedes mosquitoes. However, it is important to emphasize that the isolation of ZIKV from a mosquito is not evidence that transmission is feasible by that mosquito. Human and nonhuman primates are the main reservoirs of the virus, with humans acting as the primary host.
The most important non-vector modes of transmission include perinatal, in utero, sexual, and via blood transfusion. Although ZIKV RNA has been detected in breast milk, transmission through breastfeeding has not yet been demonstrated, reinforcing the current recommendations that mothers with ZIKV infection should continue to breast feed their infants.10
Intrauterine transmission of ZIKV, the mode with the most severe consequences, was confirmed in Brazil by the detection of virus genome, by reverse transcriptase-polymerase chain reaction (RT-PCR), in amniotic fluid samples of women with symptoms of ZIKV infection during the first trimester of pregnancy, whose fetuses had been diagnosed with microcephaly, in placental tissues from early miscarriages, and also in the blood and brain tissue of infants with congenital neurologic anomalies, including microcephaly.10,11
The virus may also be transmitted through sexual intercourse, even from asymptomatic infected individuals. Male-to-female, male-to-male and female-to-male transmissions have already been described. Interestingly, ZIKV has already been detected in the semen by RT-PCR several weeks after the infection, regardless of the presence of symptoms. In this regard, the Centers for Disease Control and Prevention (CDC) have issued preconception counseling recommending that men should use condoms for 3 months after returning from endemic areas (or after the last possible ZIKV exposure) and that women wait for 2 months before trying to conceive. Due to the ability of the ZIKV to persist in the male genital tract, there are concerns and debate on the theoretical effect of the virus on male fertility. There are few reports of possible cases of transfusion-associated transmission of ZIKV, stimulating determined countries to implement testing for ZIKV RNA in blood banks.
The incubation period in humans prior to onset of symptoms is thought to be between 3 and 14 days after the bite of an infected mosquito. Infected people, both symptomatic and asymptomatic, can transmit ZIKV to mosquitoes throughout the viremic period, that usually ranges from a few days to 1 week.10
DIAGNOSIS
Clinical diagnosis is limited by the non-specific signs and symptoms of ZIKV infection, which are similar to other arboviral infections (e.g., chikungunya and dengue), common in endemic areas. These include fever, pruritic rash, arthralgia, conjunctival hyperemia, soreness, edema of extremities, retro-orbital pain and vomiting. Abnormal laboratory findings can be present in symptomatic patients, including mild thrombocytopenia, leukopenia, and elevations in acute-phase markers of inflammation, serum lactate dehydrogenase, and/or liver transaminases.10,11
Laboratory specific tests are RT-PCR and serology – ELISA or immunofluorescence. The ZIKV RNA by RT-PCR uses serum and/or urine specimens collected within the time frame of 14 days after symptoms onset.10 ZIKV RNA can be detected during the acute viremic phase. Serum studies using ELISA or immunofluorescence can detect ZIKV-specific immunoglobulin M (IgM) and neutralizing antibodies in serum specimens collected by the end of the first week of symptoms onset. IgM can be detected as early as 5–6 days after symptoms onset, and will remain positive up to 12 weeks. As the immune response develops, IgM titers rise in peripheral blood and the level of viral RNA generally declines. Serum IgM antibody testing should be performed if the RT-PCR result is negative or when ≥14 days have passed since illness onset. IgG antibodies develop within days after IgM and can be detected for months to years. However, false-positive results due to cross-reaction with related flaviviruses (e.g., dengue and yellow fever viruses) are commonly observed. Positive results in primary flavivirus infections should be confirmed with a four-fold increase in the titer of neutralizing antibodies to ZIKV with plaque reduction neutralization test (PRNT). In endemic areas, where a great proportion of the population may have been previously infected with other flaviviruses or vaccinated against a related flavivirus (i.e., secondary flavivirus infection), neutralizing antibodies might still yield cross-reactive results in these individuals.10,11,12
CLINICAL FEATURES
It is estimated that approximately 70–80% of ZIKV infections are asymptomatic. When symptomatic, the disease is generally a mild, self-limited condition, lasting few days, and characterized by one or more of the following symptoms: low fever, pruritic rash, edema of extremities, conjunctivitis, headache, and myalgia. Less common manifestations include gastro-intestinal symptoms, retro-orbital pain and lymphadenopathy.1,10,12 Children with ZIKV infection generally have milder clinical findings when compared to adults. Although sometimes difficult to detect, arthralgia in infants and young children can manifest as irritability, limited range of motion, refusal to move one or more specific joints in the upper or lower extremities. For global standardization, WHO postulated a definition to help detect and report Zika virus cases: patients presenting with rash and/or fever, associated with at least one of the following signs and symptoms: arthralgia, arthritis or conjunctivitis (non-purulent conjunctival hyperemia). A suspected case is confirmed after lab studies showing presence of Zika virus RNA or antigen in serum or other samples – e.g. saliva, tissues, urine, whole blood; or IgM antibody against ZIKV positive, and PRNT90 for ZIKV with titer ≥20 and ZIKV PRNT90 titer ratio ≥4 compared to other flaviviruses; and exclusion of other flaviviruses.13
Neurological and auto-immune complications
Reports from several countries affected by ZIKV outbreaks list a number of neurological conditions following ZIKV infection, resulting from the neurotropism of the virus, including Guillain-Barré Syndrome (GBS), meningitis, acute disseminated encephalomyelitis and myelitis, mainly in adults.14 GBS cases following ZIKV were predominantly male individuals above the age of 60, a fact that has put ZIKV infections in the list of potential infectious pathogens that can trigger the manifestation of GBS.14,15,16
Clinical features of ZIKV infection on pregnant women
Clinical features of ZIKV infection are similar in pregnant compared to non-pregnant women, though symptomatic infections seem to be more frequent during pregnancy.17,18
Data from Brazil and Colombia showed higher rates of symptomatic ZIKV infections among women of reproductive age compared to men of same age, suggesting an increased susceptibility to ZIKV infection among women.19,20 However, it is unknown whether behavioral factors or sexual transmission mechanisms may explain this difference in the risk of ZIKV infection.
ZIKV infection and the placenta
The pathogenesis of the mother-to-child transmission of ZIKV infection is driven by the inflammation of the placenta (placentitis), including multiplication of placental macrophages (Hofbauer cells) that facilitate viral transfer from the placenta to the fetal brain. The infection of the placenta with ZIKV may lead to hypoperfusion, fetal loss, and neonatal infection. Additionally, placental macrophages or Hofbauer cells are permissive to ZIKA and facilitate viral transfer from the placenta to the fetal brain. Active replication of ZIKA in the placenta and fetal brain has been demonstrated in mice models and found in the placenta of women with fetal losses during the first and second trimester and in brain tissue of fetuses with microcephaly. ZIKV is the only vertically transmitted flavivirus that can infect cortical progenitor cells.21
ZIKV vertical transmission can occur and cause severe fetal defects. Recent estimates, based on prospective studies and case reports, have calculated a vertical transmission rate of up to 20–30%, irrespective of the trimester when maternal infection occurred. The risk of maternal-fetal transmission, fetal loss or fetal abnormalities is not related either to the presence or the severity of maternal symptoms.22
Congenital ZIKV syndrome (CZV)
The most striking consequence following the ZIKV is the one that affects fetuses and infants from infected mothers. In October 2015, several pediatricians from the Northeastern region of Brazil observed a significant increase in the number of newborns presenting with a congenital syndrome, mainly characterized by a wide range of congenital malformations, predominantly characterized by malformation of structures in the central nervous system, and associated symptoms: microcephaly (with significant cranium-facial disproportion), spasticity, convulsions, marked irritability, and brainstem dysfunction including feeding difficulties. Strong cumulative evidence confirmed the hypothesis that these infants were infected in utero. In other words, a relationship between ZIKV infection during pregnancy and these congenital abnormalities could be established. The term congenital Zika syndrome (CZS) defines the presence of severe neurological abnormalities including microcephaly, hydrocephalus, lissencephaly with agenesis of the corpus callosum, pachygyria, cerebellar dysplasia, white-matter abnormalities and multiple calcifications in the cortex and subcortical white matter. In addition, CZS includes musculoskeletal, genitourinary, craniofacial, ocular, and pulmonary manifestations1,10,23,24,25 (Figure 4).
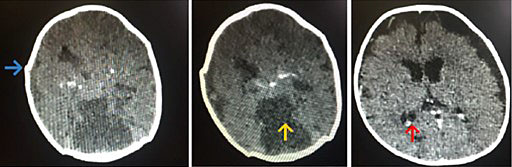
4
Computed tomographic images of infants with congenital ZIKV syndrome, showing lissencephaly, ventriculomegaly, and cerebral calcifications.
The severity of the neurological impairments seems to be related to the gestational age, i.e., the earlier gestation time at the infection onset, the more severe the neurological consequences. Other malformations described in infants with suspected congenital ZIKV infection include arthrogryposis (Figure 5), microphthalmia, funduscopic alterations in the macular region, and optic nerve abnormalities.10,23,24,25 Congenital heart malformations were also reported in a study of echocardiographic findings in infected infants, including: osteum secundum atrial septal defect, persistent foramen ovale, and minimum patent ductus arteriosus.26 The true burden of ZIKV-related congenital malformation was in fact greater than was initially estimated, with virtually all children who were born with a diagnosis of CZS presenting significant development impairments in follow-up studies. Long-term complications and sequelae were reported even in children without any clinical or radiological abnormalities at birth, that could only be identified later, when they eventually present with cognitive, developmental, and/or hearing and visual limitations. It has been estimated that up to 40% of children with prenatal ZIKV exposure may develop any type of neurodevelopmental delay.27 Furthermore, data on survival among children with congenital Zika syndrome are limited. A population-based cohort study from Brazil found mortality rate of 52.6 deaths per 1000 person-years among live-born children with congenital Zika syndrome, as compared with 5.6 deaths per 1000 person-years among those without the syndrome. The mortality rate ratio among live-born children with congenital Zika syndrome, as compared with those without the syndrome, was 11.3. These findings draw attention to the importance of primary prevention of infection in women of childbearing age against Aedes aegypti bites.28
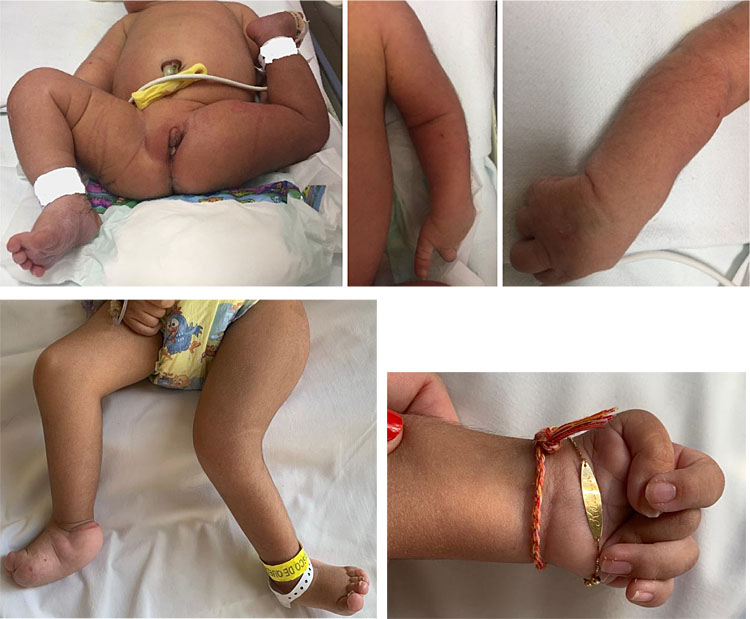
5
Infants with arthrogryposis associated with congenital ZIKV infection.
Brazil was unique in its ZIKV outbreak since the population was completely susceptible (naïve) to the virus. This led to a massive infection of highly populated urban areas from the northeast region, with a high density of Aedes aegypti. In this scenario, the linkage between in utero ZIKV infection and congenital disease was identified after the virus circulation in Brazil. Furthermore, it has been postulated that the ZIKV infection provides life-long immunity after the first infection. This reason likely explains why, in endemic places, where the virus has been circulating for years (e.g. countries in Africa and Asia), a limited proportion of childbearing age women are susceptible, due to the increased likelihood of previous exposure to the virus at younger ages. In fact, the association between ZIKV and microcephaly and other congenital malformations was only reported in the French Polynesia following the reports from Brazil that raised this causal relationship.29,30
In vitro studies showed that plasma from individuals who were previously infected with dengue virus cross-reacted with ZIKV, driving antibody-dependent enhancement of ZIKV infection.31 However, in vivo studies have not demonstrated that signs of antibody-dependent enhancement (ADE) were found in patients previously infected with dengue virus (DENV) once infected with ZIKV or vice-versa.32 In fact, a recent study performed in Brazil suggested that preexisting multitypic DENV immunity had a protective effect for the mother against congenital Zika syndrome, highlighting the complex interactions between these various Flavivirus infections.33
The devastating developmental abnormalities in infants born to women infected with ZIKV during pregnancy prompted the WHO to declare Zika a Public Health Emergency of International Concern in 2016.34 This designation spurred an intense effort to develop ZIKV vaccines.
Pregnancy loss and perinatal outcomes
ZIKV in pregnancy is also linked to several adverse pregnancy outcomes such as miscarriage, intrauterine growth restriction, stillbirths and perinatal death. While the first and second trimesters of pregnancy represent the highest fetal risk, ZIKV infections occurring later in pregnancy have also been linked with adverse outcomes including intrauterine growth restriction and fetal loss.
A systematic review found a median absolute risk of 6.3% for fetal death and 5.9% for non-fatal adverse outcomes (e.g. microcephaly).35
TREATMENT
Currently, there is no specific licensed antiviral therapy for patients with ZIKV disease. The general treatment recommendation is supportive care, including rest, fluids and symptomatic treatment (acetaminophen to relieve fever and antihistamines to treat pruritus). Aspirin and other nonsteroidal anti-inflammatory drugs (NSAIDs) should be avoided to reduce the risk of hemorrhagic complications.
Chloroquine seemed to exhibit antiviral activity against ZIKV, having reduced, in vitro, the number of ZIKV-infected cells, viral reproduction and promoting cell death, without cytotoxic effects.36 Although a number of other repurposed existing drugs (nitazoxanide, niclosamide, sofosbuvir, ribavirin, epigallocatechin catechol gallate, temoporfin, emricasan) as well as novel molecule candidates have been tested in preclinical trials (including inhibitors of ZIKV) showing potential anti-virus in vitro activity, none of them proved to show any evidence of anti-ZIKV activity in humans in clinical trials and a safe and effective drug with anti-viral activity against ZIKV is still an unmet need.37
PREVENTION
Vaccines
Preliminary studies have paved the way towards the development of an effective vaccine against ZIKV, by showing that an immune response after ZIKV infection can induce broad neutralizing antibodies (nAbs) against multiple strains of the virus (e.g. American, Asian, and early African ZIKV strains).38 ZIKV induces innate and adaptive immune response by the host. Similarly to other flaviviruses, nAbs, predominantly directed against protein E (envelope), seems to play a critical role in protection against infection. Neutralizing antibodies have been identified as a correlate of protection against ZIKV infection in mice and nonhuman primates (NHPs) and, Importantly, after transfer of IgG from human vaccines to mouse challenge models. Although the humoral response to ZIKV infection is well documented, cellular immune responses are less well understood. ZIKV-specific T CD4+ and CD8+ cell responses targeting C, E and NS1 proteins were demonstrated in humans, nonhuman primates and mice.39
The WHO Target Product Profile (TPP) for ZIKV vaccines identifies the preferred target for vaccination as women of childbearing age, potentially including pregnant women.40 The goal is to protect the fetus from infection during pregnancy, and to prevent, or at least minimize, the congenital abnormalities. Thus, it is critically important to demonstrate that vaccines not only prevent infection of adults, but that the protection induced by vaccination prior to pregnancy will prevent congenital syndrome during pregnancy. Detection of virus in semen samples collected from men several weeks after infection points to a reservoir of ZIKV that can be sexually transmitted. Thus, men are a secondary target population in the context of an emergency outbreak because of their ability to transmit virus.41
Besides, considering evidence from studies showing an increased risk of severe dengue and hospitalization among dengue-naïve subjects vaccinated with live attenuated chimeric dengue vaccine compared to the unvaccinated control group,42 it is crucial to maintain a long-term safety surveillance, to accurately assess the safety profile and the duration of the protective immune responses of current candidate ZIKV vaccines.43 It is also critical to establish a comprehensive understanding of the immune response to subsequent infections in planning for future vaccine trials. The effect of previous infections by other flaviviruses in the immune mechanisms of the vaccinated population is still unclear as is the risk of severe disease, neurological conditions like Guillain-Barré Syndrome, or congenital complications.
Different technologies have been used in the development of several candidate types of ZIKV vaccines, that can induce high levels of nAbs, as well as generating protection against the challenge of infection in non-pregnant mouse and nonhuman primate models. Inactivated virus, live attenuated virus, DNA, mRNA, viral vectors, virus-like particles (VLPs), peptide and protein-based vaccines, monoclonal antibodies (MAbs) for passive immunization, and vaccines that target vector-derived antigens are among the different platforms that have been tested for the development of a safe and effective ZIKV vaccine.44,45,46,47,48
These candidate vaccines have been tested in animal models and have been shown to be efficacious in preventing ZIKV infection of adult animals. While several of these candidates have advanced to clinical trials, it has proven challenging to clinically evaluate them for their ability to prevent congenital Zika syndrome. First, the intensity of the ZIKV epidemic has declined dramatically. This not only lessens the urgency for vaccine development, but also makes clinical endpoint studies difficult, as vaccination cohorts need to be substantially larger. Furthermore, Zika is now endemic in many areas, making it difficult to identify adequate numbers of ZIKV-naïve individuals. In addition, the low incidence of symptoms during acute disease and the low frequency of sequelae such as Guillain–Barre syndrome makes endpoints difficult to determine in clinical trials.43,49
Although several vaccine strategies have progressed into clinical trials in humans, gestational protection vaccine studies are still limited.46,47 Currently, according to ClinicalTrials.gov database, maintained by the National Library of Medicine at the National Institutes of Health (NIH), 19 phase I studies with candidate ZIKV vaccines have already been completed and three candidate vaccines progressed to phase 2 (a Zika mRNA-1893 vaccine in adult participants living in endemic and non-endemic Flavivirus areas – not yet recruiting; a purified inactivated Zika virus vaccine (PIZV) in healthy adults – not yet recruiting, and a Zika virus DNA vaccine VRC705 – completed in healthy adults and adolescents).50
DNA Vaccines
DNA vaccine platforms have been under development as candidate vaccines against several different pathogens, such as Ebola virus, Rift Valley fever virus, dengue virus, West Nile virus, chikungunya virus, and severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), for more than 25 years.47 DNA vaccines consist of plasmid DNA that contain expression promotors and the target gene, so that the target antigen is expressed in the vaccine recipient. Most currently developed DNA vaccines were constructed encoding ZIKV prM and E proteins. The DNA candidate vaccines induced both humoral and cellular immune responses, including nAbs, in mice and non-human primates. They were also capable of completely protect mice against lethal challenges of ZIKV. The Vaccine Research Center (VRC) of the National Institute of Allergy and Infectious Diseases (NIAID) and National Institutes of Health (NIH) in USA have developed two DNA vaccines expressing pre-membrane and envelope Zika virus structural proteins, one composed of a plasmid VRC5288 (Zika virus and Japanese encephalitis virus chimera), and one composed of a single plasmid VRC5283 (wild-type Zika virus), and tested them in phase I clinical trials to assess their safety, tolerability, and immunogenicity in humans.52 The Zika virus wildtype (ZIKVwt) DNA vaccine is currently being tested in a phase 2, multicenter, randomized trial to evaluate the safety, immunogenicity, and efficacy in healthy adolescents and adults.50 Another DNA-based candidate, GLS-5700, with a synthetic ZIKV prM-E sequence of multiple ZIKV isolates collected between 1952 and 2015, provided fully protection against morbidity and mortality following ZIKV challenge, decreased viral loads in blood, semen, and other tissues and conferred protection of male mice from ZIKV-induced testicular damage.51,53 Non-structural protein NS1 have also been tested as viral antigens for DNA vaccines, taking in account that functional NS1-specific T-cell responses are critical for protection against ZIKV infection. A novel DNA vaccine encoding a secreted ZIKV NS1 conferred rapid protection from systemic ZIKV infection in immunocompetent mice.54 DNA vaccines targeting the NS1 gene have the theoretical advantage of not being associated with the risk of inducing ADE in individuals from dengue and other Flavivirus endemic areas.
DNA vaccines are chemically stable and easy to produce, with no requirement for cold-chain storage, and they are also cost-effective to manufacture on a large scale. However, DNA vaccines involve expression of multiple foreign genes and thus have the potential of integrating the exogenous gene into the host genome, leading to induction of host autoimmunity.55
ZIKV mRNA vaccines
Messenger RNA (mRNA) vaccine technology has emerged as a flexible, simplified, and fast vaccine production platform. The successful experience provided by the extensive use of mRNA vaccines against COVID-19 during the pandemic has stimulated the interest in its potential use against several pathogens, including ZIKV. In particular, the real-world lessons learned from the use of mRNA vaccines in pregnant women during the COVID-19 pandemic, showing reassuring safety and effective profiles, were of paramount importance to pave the way for the use of this vaccine platform to prevent the devastating outcomes of ZIKV infection during the pregnancy for the fetus.56 Some mRNA-based vaccines against ZIKV infection, in which lipid nanoparticles encapsulate modified mRNA encoding specific ZIKV structural prM-E genes, have already been developed and tested. In animal models these vaccines induced high levels of nAbs responses that protected both immunocompetent and immunocompromised mice. Notably, a single administration of a mRNA vaccine candidate that encodes the pre-membrane and envelope (prM–E) glycoproteins of ZIKV strain Brazil SPH2015 and is encapsulated in lipid nanoparticles (LNPs) protected against a lethal dose of ZIKV, while a two-dose strategy induced strong protective immunity.57 A self-replicating mRNA vaccine platform encoding the ZIKV prM-E was developed, inducing potent humoral and cellular immune responses in mice, resulting in complete protection from ZIKV infection.58 It was also observed both in vivo and in vitro, that the risk of ADE could be avoided, maintaining protective immunity, by a modified prM-E mRNA vaccine encoding mutations, which destroyed the conserved fusion-loop epitope in the E protein.59 More recently, two phase 1 studies, performed in United States and Puerto Rico, that evaluated the safety and immunogenicity of two mRNA-based Zika virus vaccines (mRNA-1325 and mRNA-1893) in healthy adults (aged 18–49 years) who were flavivirus seronegative or flavivirus seropositive at baseline were published. The results showed that both candidates were safe and well tolerated at all evaluated dose levels. The mRNA-1325 candidate vaccine (encoding the premembrane and envelope E structural proteins (prME) from a Micronesia 2007 Zika virus isolate) elicited poor ZIKV-specific nAbs responses, while the mRNA-1893 candidate vaccine (encoding the prME from the RIO-U1 Zika virus isolate) induced strong ZIKV-specific serum nAbs responses after two doses, regardless of baseline flavivirus serostatus.60 These findings supported the continued development of mRNA-1893 candidate, that now is being evaluated in a phase 2, randomized, observer-blind, placebo-controlled, dose confirmation study to evaluate the safety, tolerability, and Immunogenicity of Zika vaccine mRNA-1893 in adults aged 18 through 65 years and living in endemic and non-endemic Flavivirus areas.50
ZIKV Vector-Based Vaccines
Viral-vector-based vaccine platforms use an antigen-encoding region genetic material from a specific pathogen inserted into the genome of a different carrier virus that, in general, has been altered to be replication incompetent and nonpathogenic.61 Adenovirus (Ad) vectors are frequently used as carriers, since they induce long-lasting humoral and cellular immune responses, are easy to engineer genetic modifications, do not require specialized cold-chain storage and are relatively inexpensive to expanded production, making them an attractive platform for production.62 Human adenovirus type 4 (Ad4), type 5 (Ad5) and type 26 (Ad26), as well as the replication-deficient chimpanzee adenoviral (ChAdOx1) have been the most extensively used vectors. Adenovirus (Ad) vectored Zika virus vaccines work by inserting a ZIKV prM-E gene expression cassette into the adenovirus carrier. Ad5-prM-E elicited both strong humoral and T-cell responses, while the Ad4-prM-E induced only a strong T-cell response after a two-dose schedule in mice.63 The humoral and cell-mediated immune responses provided by the Ad5-prM-E candidate vaccine also conferred protection against a ZIKV challenge in a mouse model. Ad26.ZIKV.M-Env, a replication-incompetent human adenovirus serotype 26 (Ad26) vector encoding the Zika membrane (M) and Env proteins based on the AdVac platform used for a vaccine to prevent Ebola virus disease and, more recently, coronavirus disease 2019 (COVID-19), induced strong and durable cellular and humoral immune responses in preclinical models, also demonstrating protective efficacy in mice and NHP. A safety and immunogenicity study performed in a non-endemic country (US) among 100 healthy adults showed that the Ad26-vectored anti-Zika virus vaccine candidate was safe and induced seroconversion in all participants 56 days after the first vaccination (GMT, 103.4 [CI, 52.7 to 202.9]), with titers persisting for at least 1 year (GMT, 90.2 [CI, 38.4 to 212.2]), as well as Env-specific cellular responses.64
Another adenovirus-based vector, the chimpanzee adenoviral vector (ChAdOx1), was also used as vaccine candidates against Zika infection. Different ChAdOx1 ZIKV vaccine candidates comprising the addition or deletion of precursor membrane (prM) and envelope, with or without its transmembrane domain (TM) were tested in pre-clinical trials. Results of these trials showed that the deletion of prM and addition of TM reduces protective efficacy and yields lower anti-envelope responses, suggesting that immunity against ZIKV can be enhanced by modulating antigen membrane anchoring, addressing important parameters in the future design of viral vectored ZIKV vaccines. Partial protection after an intravenous challenge, with reduced and delayed levels of viremia, was observed in mice that received the ChAdOx1 ZIKV vaccine candidate.65
Poxvirus-based vector platforms, such as the Modified Vaccinia Ankara (MVA), the New York Vaccinia Virus (NYVAC) and the Sementis Copenhagen Vector (SCV) representing a new vaccine vector technology based on the Copenhagen strain of Vaccinia virus, have demonstrated potential for designing novel vaccines against ZIKV. Current evidence from preclinical and clinical studies shows that these vaccine candidates can elicit both cellular and humoral immunity and provide protection in animal models.66 Ongoing clinical trials in humans are further evaluating the safety and efficacy of poxvirus-based ZIKV vaccines. A candidate ZIKV vaccine based on a highly attenuated poxvirus vector MVA expressing the ZIKV prM and E structural genes (MVA-ZIKV) induced ZIKV-specific nAbs and strong CD8+ T-cell responses. A single dose of the MVA-ZIKV vaccine reduced viremia in susceptible immunocompromised mice challenged with live ZIKV.67 Interestingly, a single-construct, multi-pathogen SCV vaccine encoding the structural gene cassettes of CHIKV and ZIKV (SCV-ZIKA/CHIK) was shown to be immunogenic and protective against both viruses in a series of mouse challenge models after a single vaccination.68
A vaccine based on the ZIKV NS1 protein from a MVA vector (MVA-ZIKV-NS1 vaccine candidate) induced strong humoral and cellular responses after a single intramuscular dose in immunocompetent mice, providing complete protection against a lethal intracerebral dose of ZIKV (strain MR766).69 The NS1 protein is a highly conserved protein, with many properties that make it an attractive target for protective antibodies. An attenuated, recombinant vesicular stomatitis virus (rVSV)-based vaccine, expressing ZIKV prM-E-NS1 with a defective methyltransferase in VSV, was highly immunogenic in mice, highlighting the important role of NS1 in ZIKV-specific cellular immune responses.70,71
ZIKV Virus-Like Particles (VLPs) Vaccines
As non-infectious entities, virus-like particles (VLPs) are empty shell structures that lack genomic material. VLPs mimic the conformation of natural viruses by expressing one or more structural proteins and thus stimulate robust antibodies in vivo. They are produced in different expression systems, such as suspension cultures of yeast, mammalian, and insect cells. Several VLP-based vaccine candidates have been developed and tested against ZIKV, demonstrating its capacity to induce robust protective immunity against ZIKV infection in mouse models.72 The delivery of VLPs, based on the use of structural proteins, can be achieved by different approaches including DNA and mRNA. Studies have shown that VLPs produced in HEK293 mammalian cells using the prM and E structural proteins, adjuvanted with aluminum, induced nAbs in animal models, providing protection against ZIKV challenge.73
A challenge in developing flavivirus vaccines is the possibility of immunogens enhancing infection through antibody-dependent enhancement (ADE). This occurs when ZIKV infection induces weakly neutralizing and cross-reactive antibodies that target the highly conserved fusion loop in EDII (EDII-FL) and may inadvertently enhance a future DENV infection, as suggested by some dose-dependent in vitro studies.74,75 Strategies to circumvent ADE are crucial in any flavivirus vaccine development. To mitigate the impact of cross-reactive ADE-promoting antibodies between ZIKV and DENV, researchers have reported several innovative mutations in or near the fusion loop of DII or DIII to reduce the production of cross-reactive antibodies. Vaccination with a Zika VLP candidate incorporating the CD loop sub-structural domain from ZIKV E protein domain III has shown immunoprotection in a mouse model of ZIKV infection. This stimulates protective antibodies associated with antibody-dependent cellular cytotoxicity (ADCC) and complement-dependent cytotoxicity (CDC) activities, offering a safer vaccine alternative to prevent ADE.76,77
ZIKV Purified Inactivated Vaccine
Inactivated virus vaccines are produced by killing the harvested virus with a chemical agent or heat. This process ensures that the virus can no longer replicate or cause disease while retaining its structural integrity, allowing it to stimulate an immune response. Inactivated vaccines are, in general, considered very safe but may have lower immunogenicity, typically necessitating multiple doses and periodic boosters. Pregnant women, who are in the greatest need for Zika vaccines, are usually included in the final stages of clinical evaluation due to concerns about potential adverse effects on the mother and developing fetus.56 However, inactivated virus vaccine platforms have a well-established safety record for both pregnant women and fetuses. A purified formalin-inactivated Zika virus vaccine (ZPIV) based on a 2015 Puerto Rican strain demonstrated protective efficacy in mice and non-human primates, preventing viremia after Zika virus challenge.78 It was safe and induced strong neutralizing antibody responses in healthy non-pregnant women and cross-protective B-cell responses against Zika and dengue viruses.79
Given their expected safety profile, inactivated virus vaccines like ZPIV adjuvanted with aluminum hydroxide are considered a potentially suitable platform for vaccinating pregnant women. Another inactivated ZPIV candidate, Takeda's TAK-426, showed good tolerability, an acceptable safety profile, and immunogenic in both flavivirus-naive and flavivirus-experienced adults. TAK-426 was selected for further clinical development based on its safety and immunogenicity profiles. A phase 2, randomized, observer-blind, placebo-controlled trial to evaluate the safety and immunogenicity of the purified inactivated Zika virus vaccine (PIZV) administered on day 1 and 29 and of a single booster dose of PIZV administered at 6 months post dose 2 in healthy subjects aged 18 to 65 years in the US is planned to start enrollment in late 2023.80 A Vero cell-adapted Zika virus strain (GMZ-002) and a purified inactivated virus (PIV) vaccine exhibited significantly increased productivity in Vero cells. IFNAR1-blocked C57BL/6 mice receiving two doses of the PIV were fully protected against lethal challenge, indicating a robust and persistent protective immunity and a promising vaccine candidate for Zika virus.81
ZIKV Live Attenuated Vaccine
Live attenuated vaccines mimic natural viral infections and typically provide rapid, effective, and long-lasting immunity, without the need of adjuvants or repeated booster vaccinations, although they may sometimes be associated with safety issues. Various candidates have been developed and tested in animal models. One such live attenuated vaccine candidate features a 10-nucleotide engineered deletion in the ZIKV genome's 3' untranslated region (30UTR 10-del ZIKV), which has demonstrated significant attenuation, immune responses, and protection in the A129 mouse model. Notably, a single dose of this attenuated vaccine induced sterilizing immunity, high levels of neutralizing antibodies, and completely prevented viremia after challenge, while also triggering a strong T-cell response in immunized mice. The attenuated 10-deletion ZIKV could not infect mosquitoes after oral feeding of spiked blood meals, adding an extra safety feature for use in non-endemic regions. The balance between immunogenicity and safety merits further development of this promising live attenuated ZIKV vaccine candidate.82 Nevertheless, ZIKV vaccine trials have excluded pregnant women due to safety concerns.
Other approaches that have been taken to create live attenuated ZIKV vaccines include engineering attenuating mutations in an authentic ZIKV isolate, generating a chimeric flavivirus using DENV, JEV, or YFV backbones to express ZIKV prM-E genes or using codon pair deoptimization.83 For attenuating mutations approach, one study showed that the attenuated ZIKV rGZ02a candidate elicited strong inhibitory antibody responses with moderate long-term durability.84 As a booster vaccine, rGZ02a significantly enhanced the protective immunity primed by Ad2-prME, an adenovirus-vectored vaccine expressing ZIKV prM and E proteins, making rGZ02a a promising live attenuated ZIKV vaccine candidate. For the chimeric approach, research indicated that chimeric DENV-2 and JEV SA14–14–2 carrying ZIKV prM-E genes protected mice from ZIKV infection after a single-dose vaccination.85,86 The JEV SA14–14–2 chimeric ZIKV vaccine also protected non-human primates from ZIKV infection and maternal-to-fetal transmission in mice. Immunization of mice and rhesus macaques with a single dose of a recombinant chimeric ZIKV vaccine candidate, ChinZIKV, which expresses the prM-E proteins and uses the licensed Japanese encephalitis live attenuated vaccine SA14–14–2 as the genetic backbone, elicited robust and long-lasting immune responses and provided complete protection against ZIKV challenge.85 Similarly, a chimeric ZIKV vaccine based on a live attenuated DENV-4 vaccine backbone (rDEN4D30) called rZIKV/D4D30–713 has been developed and was recently tested in a phase I evaluation of the safety and immunogenicity in flavivirus-naïve adults, 18–50 years in US.87
ZIKV can be attenuated using the technique of codon pair deoptimization. Three codon pair-deoptimized ZIKVs (Min E, Min NS1, and Min E+NS1) were synthesized and recovered through reverse genetics, incorporating a large number of underrepresented codon pairs in the E and/or NS1 genes. The Min E+NS1 strain elicited stronger neutralizing antibody responses after a single vaccination and subsequently safeguarded mice against lethal wild-type ZIKV infection and vertical ZIKV transmission. The codon pair deoptimization method also minimizes the risk of reverting to wild-type virulence.88
A live-attenuated Zika virus (ZIKV) vaccine was tested in a pregnant mouse model to determine its potential for preventing a congenital syndrome in fetuses of pregnant women. The results showed that maternal immunization with the vaccine did not have any adverse effects on pregnancy, fetal development, or offspring behavior. The maternal immunization protected against ZIKV infection and in utero transmission, with the transfer of maternal antibodies. It was found that higher neutralizing titers were required to protect pregnant mice against in utero transmission than that required to protect non-pregnant mice against viral infection.89
Live attenuated vaccines (LAVs) have proven to be highly effective in controlling YFV and JEV infections. Compared to other vaccine platforms, LAVs offer several advantages, including requiring only a single dose, closely resembling natural viral infection, providing fast and long-lasting protection, and being cost-effective.
ZIKV Recombinant Protein Vaccines
In recent years, computer-assisted peptide vaccines have gained considerable interest as alternative vaccine candidates. The peptide vaccine approach precisely targets the epitope region within antigens to elicit immune responses. In some cases, multiple peptides for one or more viral infections can be combined into clusters, allowing for immunization against a broad range of infections. The ZIKV E protein, as the primary target of neutralizing antibodies (nAbs) and protective immune responses, has been traditionally used in vaccine development. Some research to develop subunit vaccines for ZIKV has focused on generating recombinant ZIKV envelope protein domain III that binds to the cell surface receptor (EDIII) and the entire ectodomain (E80, comprising EDI, EDII, and EDIII) using both prokaryotic and eukaryotic expression systems as vaccine candidates. These constructs displayed immunogenicity and protective efficacy in immunocompetent mice.90 In nonhuman primates, both elicited neutralizing antibody responses, but only E80 immunization inhibited ZIKV infection in both peripheral blood and monkey tissues, whereas EDIII increased blood ZIKV RNA through possibly antibody-dependent enhancement, demonstrating the superiority of E80 as a vaccine candidate.91 These products could be further evaluated as standalone vaccine candidates or used in a heterologous prime-boost regimen with other types of ZIKV vaccines. Another strategy demonstrated that recombinant E proteins generated using baculovirus or insect cell production can induce strong immunogenicity in immunized mice.92,93 These proteins were able to inhibit ZIKV infection by triggering antigen-specific antibody and T-cell responses. Utilizing nanoadjuvants with immunostimulatory properties and delivery systems can enhance the efficacy of recombinant protein-based vaccines.94 One study showed that immunization with zEDIII together with the saponin-based nanoadjuvant IQB-80 from Q. brasiliensis significantly improved immune responses against ZIKV in mice, identifying saponin-based delivery systems as suitable adjuvants for recombinant ZIKV vaccines and providing important implications for recombinant protein-based vaccine formulations against other flaviviruses.95 While antibodies targeting the envelope glycoprotein can neutralize the virus, they pose a risk of antibody-dependent enhancement (ADE) of disease when they are subneutralizing.96 In contrast, antibodies generated against the NS1 protein can be protective without eliciting ADE, as NS1 is the most conserved antigen. In earlier studies, purified NS1 protein was investigated as a subunit vaccine candidate for numerous flaviviruses, such as YFV, DENV, and JEV. Consequently, NS1 could also be a promising vaccine candidate against ZIKV.47
Anti-ZIKV Monoclonal Antibodies
Due to the limitations on vaccine design against ZIKV infection, passive immunization using monoclonal antibodies (MAbs) has become a widely studied strategy for treating and preventing the infection. Anti-ZIKV MAbs have demonstrated the ability to neutralize infection in vitro and protect against morbidity in mouse models.97 Anti-E80 and anti-EDIII sera are particularly effective at neutralizing ZIKV in vitro, and passive transfer of these sera can protect recipient mice from lethal ZIKV challenges.98
High-throughput antibody isolation has helped improve our understanding of B-cell responses following infection or vaccination. The isolation of potent neutralizing antibodies (nAbs) and their detailed molecular examination have provided crucial insights into immunogen design and the potential for cross-flavivirus ZIKV vaccines.99
While some of the MAbs tested have shown weak responses against ZIKV and may pose a risk of developing antibody-dependent enhancement (ADE) related to ZIKV infection, monoclonal antibodies targeting the Zika virus nonstructural NS1 protein have demonstrated protection without inducing ADE. The protective efficacy of NS1-targeted MAbs may be related to their epitope recognition, and they also have the potential to suppress ZIKV pathogenicity.
Human monoclonal antibody studies have shown therapeutic efficacy in experimental models. Monoclonal antibody treatment markedly reduced tissue pathology, placental and fetal infection, and mortality in mice. Thus, nAbs can protect against maternal-fetal transmission, infection and disease, and reveal important determinants for structure-based rational vaccine design efforts.100 A phase 1, first in human, single dose ascending (6 dose cohorts) study, evaluated the safety of Tyzivumab, a ZIKV monoclonal antibody (mAb), in healthy adults aged 21–45 in Singapore. Another study planned to be conducted among ZIKV infected patients aged 21–60 years was withdrawn due to lack of eligible patients.101
Similarly, the administration of a cocktail containing three potent ZIKV-neutralizing monoclonal antibodies to nonhuman primates 1 day before challenge with ZIKV demonstrated that it completely prevented viremia in serum after challenge. Given that numerous antibodies have exceptional safety profiles in humans, the cocktail could be rapidly developed to protect uninfected pregnant women and their fetuses.102
These findings suggest that passive immunization with antibodies, particularly those targeting the NS1 protein, could be a valuable strategy for protecting against ZIKV infection.
Vector control
Prevention related to insect control relies on personal strategies to reduce vector density in endemic areas, through community-level programs. The different control methods against Aedes spp mosquitoes rely on the following categories: environmental (characterized by permanent actions to eliminate sources of Aedes spp in places with standing water); biological (including natural predators of the mosquitoes, like species of fish. The bacterium Wolbachia is also a biological method to vector control, reducing the virus replication and thus limiting transmission of the virus in transinfected mosquitoes); mechanical (different types of non-insecticidal mosquito traps, which capture the insects); chemical (insecticides); and genetic (based on the production of genetically engineered insects, which carry a dominant lethal gene capable of killing subsequent Aedes generations).103
The use of insect repellents containing DEET, picaridin, oil of lemon eucalyptus, or IR3535, as well as permethrin-treated clothing and gear are recommended to personal protection against mosquito bites.10
Although the elimination, almost 70 years ago, of A aegypti from Brazil and several other Latin-American countries was achieved after the implementation of an aggressive program of larval source reduction was implemented, it was not sustained and A aegypti eventually recolonized and expanded its range in the region. However, the past successful efforts suggest that such achievements are potentially feasible to be realized again.104
CONCLUSION
The importance of developing a vaccine to prevent congenital Zika syndrome (CZS) is emphasized by the ability of the Zika virus (ZIKV) to cross the fetal-placental barrier and cause harm to the fetus. Therefore, evaluating the efficacy of candidate vaccines in preventing these severe fetal outcomes is crucial. Several factors must be considered in this research, such as whether achieving sterilizing immunity (sych as is achieved with measles, polio, yellow fever and smallpox vaccines for example) is necessary for preventing these outcomes or if reducing infection viremia is sufficient, and if immune correlates established in non-pregnant are applicable to pregnant persons. Further research is needed to determine the impact of low-level viremia in fetal and maternal compartments. Additionally, given the differences between mice and primates, a non-human primate (NHP) pregnancy model would be preferred, as ZIKV transmission from mother to child during pregnancy has been observed in NHP, causing detrimental effects to the fetuses, particularly when infection occurred early in pregnancy. It is essential to consider the primary goal of the ZIKV vaccine in preventing CZS and to assess its protective efficacy in preclinical pregnancy models before clinical development.
According to WHO, if prevention of a virologically confirmed case of ZIKV illness, and possibly infection, is eventually not feasible to assess in a controlled clinical trial, it will need to be evaluated after licensure. It is assumed that effectiveness against the longer-term neurological complications and congenital ZIKV syndrome will only be possible to assess after authorization. It is unlikely that initial products would contain an indication for use in pregnant women at the outset. Last, but not least, it is important to acknowledge that although many vaccine candidates are under development in clinical trials, only three are on phase 2 and none of them in pregnancy.
PRACTICE RECOMMENDATIONS
- Zika virus (ZIKV) is an emerging pathogen of public health concern, transmitted primarily by Aedes mosquitoes, associated with a dramatic burden in places where the virus caused outbreaks between 2015 and 2017.
- Although reported cases of ZIKV disease declined from 2017 onwards worldwide, transmission persists at low levels in several countries in the Americas and other endemic regions, anticipating the potential risk of future outbreaks among susceptible cohorts in endemic regions.
- Symptoms are generally mild and include fever, rash, conjunctivitis, muscle and joint pain, malaise or headache. Symptoms typically last for 2–7 days. Most people with Zika virus infection do not develop symptoms.
- Zika virus infection during pregnancy can cause infants to be born with microcephaly and other congenital malformations, known as congenital Zika syndrome. Infection with Zika virus is also associated with other complications of pregnancy including preterm birth and miscarriage.
- An increased risk of neurologic complications is associated with Zika virus infection, particularly in adults but also in children, including Guillain-Barré syndrome, neuropathy and myelitis.
- There is no specific antiviral therapy for ZIKV disease. The general treatment recommendation is supportive care, including rest, fluids and symptomatic treatment.
- Interventions to reduce the burden of ZIKV infection are currently restricted to Aedes spp mosquitoes control.
- The current scenario highlights the importance of offering protection to women of childbearing age and their children by developing prophylactic maternal vaccines or interventions that are able to prevent pregnant women from developing ZIKV infection and, consequently, to prevent fetal/perinatal transmission.
CONFLICTS OF INTEREST
The author(s) of this chapter declare that they have no interests that conflict with the contents of the chapter.
Feedback
Publishers’ note: We are constantly trying to update and enhance chapters in this Series. So if you have any constructive comments about this chapter please provide them to us by selecting the "Your Feedback" link in the left-hand column.
REFERENCES
Sáfadi MAP, Nascimento-Carvalho C. Update on Zika: What You Need to Know. Pediatr Infect Dis J 2016;36(3):333–35. | |
Kuno G, Chang GJ, Tsuchiya KR, et al. Phylogeny of the genus Flavivirus. J Virol 1998;72:73–83. | |
Lin HH, Yip BS, Huang LM, et al. Zika virus structural biology and progress in vaccine development. Biotechnol Adv 2018;36(1):47–53. | |
Dick GW, Kitchen SF, Haddow AJ. Zika virus. I. Isolations and serological specificity. Trans R Soc Trop Med Hyg 1952;46:509–20. | |
Ioos S, Mallet HP, Leparc Goffart I, et al. Current Zika virus epidemiology and recent epidemics. Med Mal Infect Vol 2014;44(7):302–7. | |
WHO. ZIKA EPIDEMIOLOGY UPDATE. Available at: https://www.who.int/publications/m/item/zika-epidemiology-update--february-2022. | |
Brazil. Ministry of Health. Health Surveillance Secretariat. Boletim Epidemiológico. Monitoramento dos casos de dengue, febre de chikungunya e febre pelo vírus Zika até a Semana Epidemiológica 52 de 2022. Volume 54. 2023. Available at: https://www.gov.br/saude/pt-br/centrais-de-conteudo/publicacoes/boletins/epidemiologicos/edicoes/2023/boletim-epidemiologico-volume-54-no-01/view. | |
Secretaria de Vigilância em Saúde. Ministério da Saúde. Boletim Epidemiológico. Situação epidemiológica da síndrome congênita associada à infecção pelo vírus Zika 2015 a 2022. Volume 54. N° 1, 2023. Available at: https://www.gov.br/saude/pt-br/centrais-de-conteudo/publicacoes/boletins/epidemiologicos/edicoes/2023/boletim-epidemiologico-volume-54-no-05/view. | |
Santos LLM, de Aquino EC, Fernandes SM, et al. Dengue, chikungunya, and Zika virus infections in Latin America and the Caribbean: a systematic review. Rev Panam Salud Publica 2023;47:e34. https://doi.org/10.26633/RPSP.2023.34. | |
CDC. Zika virus. Atlanta, GA: US Department of Health and Human Services, CDC, 2021. Available at: https://www.cdc.gov/zika/hc-providers/infection-control/index.html. | |
Ioos S, Mallet HP, Leparc Goffart I, et al. Current Zika virus epidemiology and recent epidemics, Médecine et Maladies Infectieuses, 2014;44:302–7. | |
Musso D, Gubler DJ. Zika virus. Clin Microbiol Rev 2016;29:487–524. | |
World Health Organization. Zika virus disease: interim case definitions. World Health Organization, 2016. Available at: https://apps.who.int/iris/bitstream/handle/10665/204381/WHO_ZIKV_SUR_16.1_eng.pdf?sequence=1&isAllowed=y. | |
Santos T, Rodriguez A, Almiron M, et al. Zika Virus and the Guillain–Barré Syndrome – Case Series from Seven Countries. N Engl J Med 2016;375:1598–601. | |
Cao-Lormeau V-M, Blake A, Mons S, et al. Guillain-Barré syndrome outbreak associated with Zika virus infection in French Polynesia: a case control study. Lancet 2016;387:1531–9. | |
Lima MES, Bachur TPR, Aragão GF. Guillain-Barre syndrome and its correlation with dengue, Zika and chikungunya viruses infection based on a literature review of reported cases in Brazil. Acta Trop 2019;197:105064. | |
Reynolds MR, Jones AM, Petersen EE, et al. Vital Signs: Update on Zika Virus-Associated Birth Defects and Evaluation of All U.S. Infants with Congenital Zika Virus Exposure – U.S. Zika Pregnancy Registry, 2016. MMWR Morb Mortal Wkly Rep 2017;66:366–73. | |
Marbán-Castro Elena, Goncé Anna, Fumadó Victoria, et al. Zika virus infection in pregnant travellers and impact on childhood neurodevelopment in the first two years of life: a prospective observational study. Travel Med Infect Dis 2021;40:101985. | |
Coelho Flavio Codeço, Durovni Betina, Saraceni Valeria, et al. Higher incidence of Zika in adult women than adult men in Rio de Janeiro suggests a significant contribution of sexual transmission from men to women. Int J Infect Dis IJID Off Publ Int Soc Infect Dis 2016;51:128–32. | |
Pacheco Oscar, Beltrán Mauricio, Nelson Christina A, et al. Zika virus disease in Colombia – Preliminary report. N Engl J Med 2020;383(6):e44. | |
Evans-Gilbert T. Vertically transmitted chikungunya, Zika and dengue virus infections: The pathogenesis from mother to fetus and the implications of co-infections and vaccine development. Int J Pediatr Adolesc Med 2020;7(3):107–11. | |
Pomar Léo, Musso Didier, Malinger Gustavo, et al. Zika virus during pregnancy: From maternal exposure to congenital Zika virus syndrome. Prenat Diagn 2019;39(6):420–30. | |
European Center for Disease Prevention and Control. 2016. Zika virus disease epidemic: potential association with microcephaly and Guillain-Barré syndrome. ECDC, Stockholm, Sweden. http://ecdc.europa.eu/en/publications/Publications/rapid-risk-assessment-zika-virus-first-update-jan-2016.pdf. | |
França GA, Schuler-Faccini L, Oliveira WK, et al. Congenital Zika virus syndrome in Brazil: a case series of the first 1501 livebirths with complete investigation. Lancet 2016. | |
Mlakar J, Korva M, Tul N, et al. Zika virus associated with microcephaly. N Engl J Med 2016;374:951–8. | |
Cavalcanti DD, Alves LV, Furtado GJ, et al. Echocardiographic findings in infants with presumed congenital Zika syndrome: Retrospective case series study. PLoS ONE 2017;12(4):e0175065. | |
Mulkey Sarah B, Arroyave-Wessel Margarita, Peyton Colleen, et al. Neurodevelopmental abnormalities in children with in utero Zika virus exposure without congenital Zika syndrome. JAMA Pediatr 2020;174(3):269. | |
Paixao ES, Cardim LL, Costa MCN, et al. Mortality from Congenital Zika Syndrome – Nationwide Cohort Study in Brazil. N Engl J Med 2022;386(8):757–67. | |
Brasil P, Pereira Jr JP, Gabaglia C, et al. Zika virus infection in pregnant women in Rio de Janeiro – Preliminary report. N Engl J Med 2016;16:742–52. | |
Cauchemez S, Besnard M, Bompard P, et al. Association between Zika virus and microcephaly in French Polynesia, 2013–15: a retrospective study. Lancet 2016;387:2125–32. | |
Dejnirattisai W, Supasa P, Wongwiwat W, et al. Dengue virus sero-cross-reactivity drives antibody-dependent enhancement of infection with Zika virus. Nat Immunol 2016;17:1102–8. | |
Terzian ACB, Schanoski AS, Mota MTO, et al. Viral Load and Cytokine Response Profile Does Not Support Antibody-Dependent Enhancement in Dengue-Primed Zika Virus-Infected Patients. Clin Infect Dis 2017;65(8):1260–5. | |
Pedroso C, Fischer C, Feldmann M, et al. Cross-protection of dengue virus infection against congenital Zika syndrome, Northeastern Brazil. Emerg Infect Dis 2019;25:1485–93. | |
World Health Organization. Zika situation report. Zika and potencial complications. 2016. https://www.who.int/emergencies/zika-virus/situation-report/who-zika-situation-report-12-02-2016.pdf. | |
Leisher Susannah Hopkins, Balalian Arin A, Reinebrant Hanna, et al. Systematic review: fetal death reporting and risk in Zika-affected pregnancies. Trop Med Int Health 2021;26(2):133–45. | |
Delvecchio R, Higa L, Pezzuto P, et al. Chloroquine inhibits Zika Virus infection in different cellular models. Viruses 2016;8(12). | |
Bernatchez JA, Tran LT, Li J, et al. Drugs for the Treatment of Zika Virus Infection. J Med Chem 2020;63(2):470–89. | |
Dowd KA, DeMaso CR, Pelc RS, et al. Broadly Neutralizing Activity of Zika Virus-Immune Sera Identifies a Single Viral Serotype. Cell Rep 2016;16(6):1485–91. | |
Pardy R, Rajah MM, Condotta SA, et al. Analysis of the T Cell Response to Zika Virus and Identification of a Novel CD8+ T Cell Epitope in Immunocompetent Mice. PLoS Pathog 2017;13:e1006184. | |
Vannice KS, Giersing BK, Kaslow DC, et al. Meeting Report: WHO consultation on considerations for regulatory expectations of Zika virus vaccines for use during an emergency. Vaccine 2016. World Health Organization. WHO Target Product Profiles (TPPs). https://www.who.int/immunization/research/ppc-tpp/target_product_profiles/en/. | |
da Silva LRC. Zika Virus Trafficking and Interactions in the Human Male Reproductive Tract. Pathogens 2018;7(2):51. doi: 10.3390/pathogens7020051. | |
Halstead SB. Safety issues from a Phase 3 clinical trial of a live-attenuated chimeric yellow fever tetravalent dengue vaccine. Hum Vaccin Immunother 2018;14(9):2158–62. | |
Kim IJ, Blackman MA, Lin JS. Pre-Clinical Pregnancy Models for Evaluating Zika Vaccines. Trop Med Infect Dis 2019;4(2):58. | |
Abbink P, Larocca R, De La Barrera R, et al. Protective efficacy of multiple vaccine platforms against Zika virus challenge in rhesus monkeys. Science 2016;353(6304):1129–32. | |
Poland G, Kennedy R, Ovsyannikova I, et al. Development of vaccines against Zika virus. Lancet Infect Dis 2017. http://dx.doi.org/10.1016/S1473-3099(18)30063-X. | |
Pattnaik A, Sahoo BR, Pattnaik AK. Current Status of Zika Virus Vaccines: Successes and Challenges. Vaccines (Basel) 2020;8(2):266. | |
Wang Y, Ling L, Zhang Z, et al. Current Advances in Zika Vaccine Development. Vaccines 2022;10:1816. | |
Zhou K, Li C, Shi W, et al. Current Progress in the Development of Zika Virus Vaccines. Vaccines 2021;9:1004. | |
Sáfadi MAP, Almeida FJ. The Zika Virus Epidemic Four Years After: Where are We? Am J Perinatol 2019;36(S 02):S4–8. doi: 10.1055/s-0039-1691769. | |
ClinicalTrials.gov. U.S. Department of Health and Human Services, National Institutes of Health, National Library of Medicine, and National Center for Biotechnology Information. Available at: https://clinicaltrials.gov/ct2/show/NCT03443830?term=vaccine&cond=zika+AND+%22Zika+Virus+Infection%22&draw=3&rank=20. | |
Tebas P, Roberts CC, Muthumani K, et al. Safety and immunogenicity of an anti-zika virus DNA vaccine. N Engl J Med 2021;385:e. | |
Gaudinski MR, Houser KV, Morabito KM, et al. Safety, tolerability, and immunogenicity of two zika virus DNA vaccine candidates in healthy adults: Randomised, open-label, phase 1 clinical trials. Lancet 2018;391:552–62. | |
Kudchodkar SB, Choi H, Reuschel EL, et al. Rapid response to an emerging infectious disease – lessons learned from development of a synthetic DNA vaccine targeting zika virus. Microbes Infect 2018;20;676–84. | |
Grubor-Bauk B, Wijesundara DK, Masavuli M, et al. Ns1 DNA vaccination protects against zika infection through t cell-mediated immunity in immunocompetent mice. Sci Adv 2019;5:eaax2388. | |
Zhang N, Zheng BJ, Lu L, et al. Advancements in the development of subunit influenza vaccines. Microbes Infect 2015;17:123–34. | |
Safadi MAP, Spinardi J, Swerdlow D, et al. COVID-19 disease and vaccination in pregnant and lactating women. Am J Reprod Immunol 2022;88(1):e13550. doi: 10.1111/aji.13550. | |
Medina-Magües LG, Gergen J, Jasny E, et al. mRNA Vaccine Protects against Zika Virus. Vaccines 2021;9:1464. | |
Zhong Z, Portela Catani JP, McCafferty S, et al. Immunogenicity and protection efficacy of a naked self-replicating mRNA-based Zika virus vaccine. Vaccines 2019;7:96. | |
Richner JM, Himansu S, Dowd KA, et al. Modified mRNA vaccines protect against Zika virus infection. Cell 2017;169:176. | |
Essink B, Chu L, Seger W, et al. The safety and immunogenicity of two Zika virus mRNA vaccine candidates in healthy flavivirus baseline seropositive and seronegative adults: the results of two randomised, placebo-controlled, dose-ranging, phase 1 clinical trials. Lancet Infect Dis 2023 (published online Jan 19). https://doi.org/10.1016/S1473-3099(22)00764-2. | |
Garcia G Jr, Chakravarty N, Abu AE, et al. Replication-Deficient Zika Vector-Based Vaccine Provides Maternal and Fetal Protection in Mouse Model. Microbiol Spectr 2022;10(5):e0113722. doi: 10.1128/spectrum.01137-22. Epub 2022 Sep 28. | |
Coughlan L, Kremer EJ, Shayakhmetov DM. Adenovirus-based vaccines-a platform for pandemic preparedness against emerging viral pathogens. Mol Ther 2022. | |
Bullard BL, Corder BN, Gorman MJ, et al. Efficacy of a T cell-biased adenovirus vector as a zika virus vaccine. Sci Rep 2018;8:18017. | |
Salisch NC, Stephenson KE, Williams K, et al. A Double-Blind, Randomized, Placebo-Controlled Phase 1 Study of Ad26.ZIKV.001, an Ad26-Vectored Anti-Zika Virus Vaccine. Ann Intern Med 2021;174(5):585–94. | |
Lopez-Camacho C, De Lorenzo G, Slon-Campos JL, et al. Immunogenicity and efficacy of zika virus envelope domain iii in DNA, protein, and chadox1 adenoviral-vectored vaccines. Vaccines 2020;8:307. | |
Prow NA, Jimenez Martinez R, Hayball JD, et al. Poxvirus-based vector systems and the potential for multi-valent and multi-pathogen vaccines. Expert Rev Vaccines 2018;17:925–34. | |
Perez P, Q Marín M, Lazaro-Frias A, et al. A vaccine based on a modified vaccinia virus ankara vector expressing zika virus structural proteins controls zika virus replication in mice. Sci Rep 2018;8:17385. | |
Prow NA, et al. A vaccinia-based single vector construct multi-pathogen vaccine protects against both Zika and chikungunya viruses. Nat Commun 2018;9:1230. | |
Brault AC, Domi A, McDonald EM, et al. A zika vaccine targeting ns1 protein protects immunocompetent adult mice in a lethal challenge model. Sci Rep 2017;7:14769. | |
Li A, Yu J, Lu M, et al. A zika virus vaccine expressing premembrane-envelope-ns1 polyprotein. Nat Commun 2018;9:3067. | |
Carpio KL, Barrett ADT. Flavivirus NS1 and Its Potential in Vaccine Development. Vaccines 2021;9:622. | |
Elong Ngono A, Vizcarra EA, Tang WW, et al. Mapping and role of the cd8(+) t cell response during primary zika virus infection in mice. Cell Host Microbe 2017;21:35–46. | |
Vang L, Morello CS, Mendy J, et al. Zika virus-like particle vaccine protects ag129 mice and rhesus macaques against zika virus. PLoS Negl Trop Dis 2021;15:e0009195. | |
Dejnirattisai W, Supasa P, Wongwiwat W, et al. Dengue virus sero-cross-reactivity drives antibody-dependent enhancement of infection with zika virus. Nat Immunol 2016;17:1102–8. [CrossRef]. | |
Stettler K, Beltramello M, Espinosa DA, et al. Specificity, cross-reactivity, and function of antibodies elicited by zika virus infection. Science 2016;353:823–6. | |
Lin HH, Yang SP, Tsai MJ, et al. Dengue and zika virus domain iii-flagellin fusion and glycan-masking e antigen for prime-boost immunization. Theranostics 2019;9:4811–26. [CrossRef] [PubMed]. | |
Cimica V, Williams S, Adams-Fish D, et al. Zika virus-like particle (vlp) vaccine displaying envelope (e) protein cd loop antigen elicits protective and specific immune response in a murine model. Biochem Biophys Res Commun 2020;529:805–11. | |
Faria NR, Azevedo R, Kraemer MUG, et al. Zika virus in the americas: Early epidemiological and genetic findings. Science 2016;352:345–9. [CrossRef] [PubMed]. | |
Stephenson KE, Walsh SR, Walsh SR, et al. Safety and immunogenicity of a zika purified inactivated virus vaccine given via standard, accelerated, or shortened schedules: A single-centre, double-blind, sequential-group, randomised, placebo-controlled, phase 1 trial. Lancet Infect Dis 2020;20:1061–70. | |
WHO. WHO Vaccine Pipeline Tracker. international clinical trials registry platforms – https://trialsearch.who.int/. accessed on April 2023. | |
Oh HS, Yoon JW, Lee S, et al. A purified inactivated vaccine derived from vero cell-adapted zika virus elicits protection in mice. Virology 2021;560:124–30. | |
Shan C, Muruato AE, Nunes BTD, et al. A live-attenuated zika virus vaccine candidate induces sterilizing immunity in mouse models. Nat Med 2017. | |
Adam A, Lee C, Wang T. Rational Development of Live-Attenuated Zika Virus Vaccines. Pathogens 2023;12:194. | |
Ye X, Liu X, Shu T, et al. A live-attenuated zika virus vaccine with high production capacity confers effective protection in neonatal mice. J Virol 2021;95:e0038321. | |
Li XF, Dong HL, Wang HJ, et al. Development of a chimeric zika vaccine using a licensed live-attenuated flavivirus vaccine as backbone. Nat Commun 2018;9:673. | |
Xie X, Yang Y, Muruato AE, et al. Understanding zika virus stability and developing a chimeric vaccine through functional analysis. MBio 2. | |
WHO International Clinical Trials Registry Platforms. Accessed at https://trialsearch.who.int/Trial2.aspx?TrialID=NCT03611946. | |
Li P, Ke X, Wang T, et al. Zika Virus Attenuation by Codon Pair Deoptimization Induces Sterilizing Immunity in Mouse Models. J Virol 2018;92:e00701–18. | |
Shan C, Xie X, Luo H, et al. Maternal vaccination and protective immunity against Zika virus vertical transmission. Nat Commun 2019;10(1):5677. | |
Liang H, Yang R, Liu Z, et al. Recombinant zika virus envelope protein elicited protective immunity against zika virus in immunocompetent mice. PLoS ONE 2018;13:e0194860. | |
Yang R, Liu Q, Pang W, et al. Two immunogenic recombinant protein vaccine candidates showed disparate protective efficacy against Zika virus infection in rhesus macaques. Vaccine 2021;39(6):915–25. | |
Zhu B, Ye J, Lu P, et al. Induction of antigen-specific immune responses in mice by recombinant baculovirus expressing premembrane and envelope proteins of west nile virus. Virol J 2012;9:132. | |
Qu P, Zhang W, Li D, et al. Insect cell-produced recombinant protein subunit vaccines protect against zika virus infection. Antiviral Res 2018;154:97–103. | |
Lu L, Duong VT, Shalash AO, et al. Chemical conjugation strategies for the development of protein-based subunit nanovaccines. Vaccines 2021;9:563. | |
Cibulski S, Varela APM, Teixeira TF, et al. Zika virus envelope domain iii recombinant protein delivered with saponin-based nanoadjuvant from quillaja brasiliensis enhances anti-zika immune responses, including neutralizing antibodies and splenocyte proliferation. Front Immunol 2021;12:632714. | |
Halstead SB. Neutralization and antibody-dependent enhancement of dengue viruses. Adv Virus Res 2003;60:421–67. doi: 10.1016/s0065-3527(03)60011-4. | |
Kudchodkar SB, Choi H, Reuschel EL, et al. Rapid response to an emerging infectious disease – lessons learned from development of a synthetic DNA vaccine targeting zika virus. Microbes Infect 2018;20:676–84. | |
Qu P, Zhang W, Li D, et al. Insect cell-produced recombinant protein subunit vaccines protect against zika virus infection. Antiviral Res 2018;154:97–103. | |
Dussupt V, Modjarrad K, Krebs SJ. Landscape of monoclonal antibodies targeting zika and dengue: Therapeutic solutions and critical insights for vaccine development. Front Immunol 2020;11:621043. | |
Sapparapu G, Fernandez E, Kose N, et al. Neutralizing human antibodies prevent Zika virus replication and fetal disease in mice. Nature 2016;540:443–7. doi: 10.1038/nature20564. | |
ClinicalTrials.govBETA. U.S. Department of Health and Human Services, National Institutes of Health, National Library of Medicine, and National Center for Biotechnology Information. Available at: https://beta.clinicaltrials.gov/study/NCT03443830?distance=50&cond=Zika%20Virus&term=Monoclonal%20Antibody&rank=1. | |
Magnani DM, Rogers TF, Beutler N, et al. Neutralizing human monoclonal antibodies prevent Zika virus infection in macaques. Sci Transl Med 2017;9:eaan8184. doi: 10.1126/scitranslmed.aan8184. | |
Vanlerberghe V, Gómez-Dantés H, Vazquez-Prokopec G, et al. Changing paradigms in Aedes control: considering the spatial heterogeneity of dengue transmission. Rev Panam Salud Publica 2017;41:e16. | |
Webb JLA. Aedes aegypti suppression in the Americas: historical perspectives. The Lancet 2016;388(10044):556–7. doi: 10.1016/s0140-6736(16)31225-9. |
Online Study Assessment Option
All readers who are qualified doctors or allied medical professionals can automatically receive 2 Continuing Professional Development points plus a Study Completion Certificate from GLOWM for successfully answering four multiple-choice questions (randomly selected) based on the study of this chapter. Medical students can receive the Study Completion Certificate only.
(To find out more about the Continuing Professional Development awards program CLICK HERE)