Intracytoplasmic Sperm Injection and Other Micromanipulation Techniques for Assisted Reproduction
Authors
INTRODUCTION
Microsurgical techniques have been used in studies of living cells for over a century, with the first documented microdissection of a protozoan dating back to 1835, some 200 years after the microscope became a useful tool for scientific investigations.1, 2 These techniques are usually performed on single cells or a small group of cells under a microscope to alter their structure or composition, and are collectively referred to as micromanipulation. In the past few decades micromanipulation techniques have aided many scientific advancements in numerous disciplines. For example, cloning and genetic engineering of animals for research were made possible by micromanipulation of mammalian eggs and embryos. In the 1980s, micromanipulation of human gametes and embryos began to improve the efficacy of assisted reproductive technologies (ART) to treat infertility, and has since become an integral part of ART. Micromanipulation is an indispensable tool in other medical procedures derived from ART, such as genetic testing of in vitro generated embryos. This chapter reviews established and new applications of micromanipulation techniques in ART, with an emphasis on our understanding of the benefits and risks of these techniques. More comprehensive, historical reviews of this subject can be found elsewhere.3, 4, 5, 6, 7
MICROASSISTED FERTILIZATION
In the conventional in vitro fertilization (IVF) procedure, oocytes are coincubated with motile sperm to achieve fertilization. This conventional method usually produces a fertilization rate averaging between 50% and 70%. In the case of infertility caused by a male factor, fertilization can be severely compromised, if it occurs at all. Improving fertilization for male infertility was probably the first application of micromanipulation in ART.
Human oocytes, similar to oocytes from most mammalian species, are covered by a transparent glycoprotein coat called the zona pellucida (ZP). In order to gain access to the oocyte plasma membrane (oolemma) for fertilization, a sperm has to be able to negotiate its way through this barrier. Microassisted fertilization techniques have been designed to circumvent this ZP barrier. The first technique reported for human IVF, known as zona drilling, involved “driling” a hole in the ZP by applying an acidic solution (e.g., Tyrode’s solution with pH adjusted to 2–3) to a localized area with a micropipette.8 Partial zona dissection (PZD) refers to a micromanipulation procedure whereby the ZP is sliced open by a microneedle.9 Zona opening can also be accomplished by laser ablation.10 An alternative to zona opening is to place sperm directly next to the oolemma in the perivitelline space using a micropipette, which is named SUZI (subzonal insemination).11 The efficacy of these assisted fertilization techniques is compromised by two problems. First, not all sperm can effectively interact with the oolemma for the subsequent fertilization events to occur, even after they have bypassed the ZP barrier. Second, the block to polyspermic fertilization is not effective at the oolemma level in the human. Thus, multiple sperm could enter the oocyte resulting in polyploidy. Consequently, these techniques have produced relatively low and inconsistent fertilization results as treatment for male infertility. However, these techniques have the advantages of being relatively atraumatic to the oocyte and allowing, to some extent, natural selection of the fertilizing sperm.
The injection of a single sperm into the cytoplasm of the oocyte, or intracytoplasmic sperm injection (ICSI), provided a satisfactory solution to the problems of the assisted fertilization techniques developed earlier.12 In this procedure, a single sperm is first immobilized by touching the sperm tail with an injection pipette (inner diameter 5–7 μm). The injection pipette picks up the immobilized sperm, pierces the ZP and oolemma, and delivers the sperm inside the oocyte cytoplasm (Fig. 1). In 1976 using hamsters as a model, Uehara and Yanagimachi13 were probably the first to report the injection of sperm into oocyte cytoplasm (ooplasm). It was later attempted on rabbit14 and human oocytes,15 although the first successful human pregnancy was not reported until 1992 by the Free University of Brussels’ group in Belgium.12 During the natural fertilization process, the human sperm undergoes acrosome reaction on binding to ZP sperm receptor. The acrosome reaction enables the sperm to penetrate the ZP and to come into contact with the oolemma. The sperm cell membrane fuses with the oolemma and the sperm cell is incorporated into the ooplasm. The fusion activates the oocyte. Only after activation can an oocyte proceed to complete meiosis with the extrusion of the second polar body and allow sperm nucleus decondensation.16 In comparison with this natural process, ICSI does not require the sperm to complete the acrosome reaction. Fusion between sperm and oocyte membranes does not take place. Instead, the dislodging of the ooplasm by the ICSI pipette serves as a trigger for oocyte activation.17, 18, 19 The breakage of cell membrane on the sperm tail during immobilization facilitates the release of a putative oocyte-activation factor from the sperm that is also essential for activating the oocyte.20, 21
|
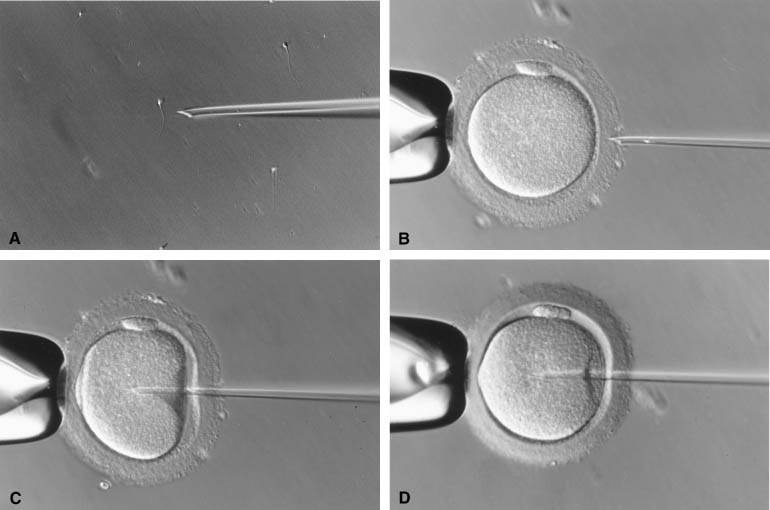
Because ICSI is much more effective in fertilizing oocytes than other assisted fertilization techniques, it has become the ART of choice for male infertility.5, 22, 23 Indications for ICSI encompass most types of male infertility. Satisfactory fertilization can be achieved by ICSI when any of the following is diagnosed: low sperm count or motility, poor sperm morphology, presence of antisperm antibodies in the male (auto-) or female (allo-), or prior experience of failed or suboptimal fertilization with conventional IVF. In case of azoospermia or necrospermia, viable sperm can be extracted surgically from the epididymis or testis under many circumstances, and such surgically collected sperm can effectively fertilize oocytes with the aid of ICSI.24, 25, 26 Cryopreservation does not significantly reduce the fertilizing ability of surgically collected sperm,27, 28, 29 which negates having to schedule the urology surgery and oocyte retrieval on the same day. Although immature spermatozoa have been successfully used for fertilization by ICSI,30 treatment outcome after using spermatids has been disappointing, largely because of the difficulty in correctly identifying round spermatids from somatic cells.31, 32 For semen sample with poor sperm motility, fertilization may be further enhanced by ICSI after treating the sperm with phosphodiesterase inhibitors, such as pentoxifylline.33, 34
The applications of ICSI have expanded beyond treating male infertility. ICSI can be used when suboptimal fertilization is anticipated because of the adverse effects of some in vitro conditions. For example, mammalian oocytes matured in vitro tend to develop a hardened ZP (resistance to proteinase digestion) that may hinder fertilization.35, 36 Optimal fertilization of in vitro matured oocytes has been obtained by ICSI.37, 38, 39 Similarly, cryopreservation also causes zona hardening,40, 41 and fertilization of the cryopreserved human oocytes can be optimized by the use of ICSI procedure.42, 43 The combination of ovarian tissue cryopreservation followed by in vitro oocyte maturation has become one of the potential avenues for fertility preservation prior to cancer therapy in women of reproductive age.44 Fertilization of such oocytes is ensured by ICSI.45 In the conventional IVF method, oocytes are coincubated with a suspension of 50,000–100,000 sperm for fertilization. In comparison, oocytes are exposed to individual sperm during ICSI. Thus, IVF by ICSI can greatly reduce the likelihood of transmitting microorganisms from the male to the female partner. Successful attempts have been reported for using ICSI as a procreation method to prevent transmission of human immune deficiency viruses between couples with discordant serotypes.46, 47 However, this practice has been cautioned against by the findings from rhesus monkey studies in which foreign DNA attached to sperm was introduced to oocytes via ICSI and the DNA was subsequently incorporated as functional genes into the embryo genome.48, 49 Taken together, these studies suggest that ICSI may minimize, but does not eliminate, the chance of transmitting infectious diseases between partners. The procedure may facilitate the introduction of foreign DNA and other pathogens to the oocytes, potentially leading to greater health complications. In some ART centers, the conventional IVF method has been replaced by ICSI not only for male infertility but also for infertility without an identified cause (unexplained infertility) in order to ensure that fertilization occurs. This is because fertilization usually fails to occur in 2–5% of IVF attempts even though all semen parameters are within normal ranges. The use of ICSI may prevent such complete failures.50 However, fertilization failure may still occur even when ICSI is used.51 Therefore, taking into account the added expenses and the potential risks of the procedure, it remains debatable whether ICSI should be used exclusively in place of the conventional IVF method for all patients.52
As ICSI becomes an established ART procedure, many concerns have been raised over its potential detrimental effects on the resultant embryos and children. One of the main concerns arises from the fact that ICSI bypasses the natural selection of sperm for fertilization. Sperm with defects that would have prevented them from fertilizing ooyctes may do so with the aid of ICSI and may pass the defects on to the next generation. Submicroscopic deletions (microdeletions) in the long arm of the Y chromosome have been found in approximately one fifth of men with azoospermia or severe oligospermia who would be prime candidates for ICSI.53, 54, 55 These microdeletions are probably de novo events and may be transmitted to the male offspring.56 In some men the microdeletions are caused by mitotic errors postfertilization, resulting in mosaicism in the affected individual. For these men, microdeletions may not be transmitted to all male offspring.57 Furthermore, because a direct cause relation has not been established between these chromosomal anomalies and fertility, it is uncertain whether the transmission of Y chromosome microdeletions will necessarily render all the male offspring infertile. Sperm from infertile men have a higher prevalence of aneuploidy than those from fertile controls.58, 59 This may explain, at least partially, the small increase in the incidence of chromosomal anomalies in ICSI-generated embryos.60, 61, 62
Apart from facilitating the transmission of genetic defects, the process of sperm injection may inevitably cause physical damages to the oocyte and interfere with subsequent embryo development. In comparison with embryos from the conventional IVF method, ICSI-generated embryos have been found to be less likely to attain the blastocyst stage in vitro and more likely to develop fragments.63, 64, 65 However, such differences were not observed in other studies.66, 67, 68 The discrepancy may be partially the result of technical differences or operator variations, which are known to influence fertilization and embryo development.69, 70 Because the position of the second meiotic spindle is variable and is not always beneath the first polar body,71, 72 the insertion of ICSI pipette could damage the meiotic spindle even if the area adjacent to the first polar body is avoided. Wang and coworkers73 reported a polarizing optical system that allows the visualization of the meiotic spindle without destroying the oocyte. Under the guidance of this system, damage to the spindle by the ICSI pipette can be avoided, thereby reducing the incidence of chromosomal anomalies or other cellular damages in ICSI-generated embryos. To prepare for the ICSI procedure, motile sperm are usually placed in a high-viscosity medium containing 10% (w/v) of a synthetic polymer, polyvinylpyrrolidone (PVP). A small amount of PVP is inevitably injected into the oocyte together with the sperm. To date, there is no evidence that PVP has any detrimental effect on oocytes or embryos. Nevertheless, the replacement of PVP with a natural material, hyaluronic acid, has been reported for preparing sperm for ICSI but its benefit remains to be confirmed.74
Despite the observed and potential detrimental effects on the embryos at the genetic and cellular levels, ICSI has not been associated with increased incidence of birth defects.75, 76, 77, 78 Recently, public concerns over the safety of ART were raised again by a report showing a higher incidence of birth defects in IVF babies.79 In that report, however, the incidence of birth defects was not further increased by ICSI. It is possible that the damage done by ICSI, if any, is restricted to early stages of embryo development. Embryos that survive this early stage do not have further disadvantages compared to embryos resulting from conventional IVF during their subsequent development. The limited information available suggests that ICSI children may have a small delay in mental development,80, 81 although it is unknown if this mental impairment is caused by the ICSI procedure or by factors inherent to the patients who require ICSI in the first place. Concerns were raised when imprinting disorders were found in a small number of children born from ICSI procedures.82, 83 However, closer examination of large numbers of children born after fertility treatments, including ICSI, dimissed any direct association between ICSI and the risk of imprinting defects.84, 85
Obviously further studies of the long-term effects of ICSI on the offspring involving multiple centers with well-controlled study designs are needed to minimize confounding variables, such as operator/technical variations and population variations. Until conclusive data become available, patients should be counseled carefully before ICSI is offered as an ART treatment.
MICROASSISTED IMPLANTATION
Embryo implantation has long been recognized as the bottleneck limiting the success of ART treatments. Unfortunately, little is known of the pathophysiology underlying implantation failures after ART. Scientific research in this area has been hampered not only by technical difficulties but also by ethical complications because scientific research would inevitably have to involve experimentation on human embryos. Despite the lack of knowledge of the fundamentals of embryo implantation, new technologies have been developed in the past decade to improve the implantation potential of in vitro-produced embryos.
Assisted hatching
Assisted hatching is a laboratory procedure developed to improve the implantation potential of in vitro-produced embryos by creating an opening in the ZP.86, 87 The ZP persists after fertilization until the embryo is ready to implant in the uterus. Under in vivo conditions, hatching is a misnomer for most mammalian species. The ZP of mammalian embryos is not cracked open from the inside by the embryo, rather, it is dissolved from outside by proteolytic agents in the uterine fluid.88, 89, 90, 91, 92 Only under in vitro conditions is the embryo observed to generate an opening in the ZP through which the embryo escapes, leaving behind an empty zona. If assisted hatching can benefit embryo implantation, it probably does so primarily by assisting the uterus, rather than the embryo, in removing the ZP. However, this is not to imply that the embryo has nothing to do with the zona loss in vivo. The loss of ZP is most likely a result of embryo-uterine interactions. For example, the embryo may secrete a protease activator that activates a latent protease in the uterine fluid, and the activated uterine protease then acts to digest the ZP.93 Moreover, certain zona characteristics, such as thickness and resistance to enzyme digestion, have been found to correlate with embryo quality and implantation potential.94, 95, 96, 97
Assisted hatching can be accomplished by the same methods as those developed initially for assisted fertilization, such as zona drilling, PZD, and zona ablation with laser.8, 9, 10 During the zona drilling procedure, the same micropipette that delivers the acid can be used to remove fragments from the embryo. The presence of fragments in IVF-generated human embryo is not uncommon and may interfere with cell-cell interactions within the embryo. Therefore, the removal of fragments can presumably further improve embryo development. When using acid for assisted hatching, it is important to avoid acid contact with embryonic cells. Once a hole is made, excessive acid solution near the hatched site should be removed by the hatching pipette and the embryo should be rinsed in fresh media immediately following the hatching procedure (Fig. 2). In PZD, cross slits have been shown to be more effective than the originally described two-dimensional slicing method.98 To mimic more closely the in vivo events of zona loss, the ZP can be globally thinned by laser or completely dissolved enzymetically or chemically.99, 100, 101, 102 A study evaluated four different assisted hatching methods (zona drilling, PZD, laser ablation, and enzymatic thinning) and found that these different methods produced similar implantation and pregnancy rates.103
|

Although assisted hatching has been practiced in many ART centers for some time, it remains inconclusive whether assisted hatching can improve ART treatment outcome. Some studies support the initial observation by Cohen and colleagues104 that assisted hatching improves implantation and pregnancy rates;105, 106, 107, 108 while other studies disagree.109, 110, 111, 112 These conflicting observations are difficult to reconcile because of many important differences between the studies, such as study design, patient population, sample size, and assisted hatching techniques or skills. It is likely that assisted hatching only benefits a selective group of patients, provided that the procedure is performed correctly by an experienced operator. In the Practice Committee Report in 2000, the American Society for Reproductive Medicine suggests that “assisted hatching may be clinically useful” and recommends that “individual ART programs should evaluate their own patient populations in order to determine which subgroups may benefit from the procedure. The routine or universal performance of assisted hatching in the treatment of all IVF patients appears, at this point, to be unwarranted.”113 In spite of its uncertain benefits, assisted hatching has not been associated with impaired embryo development or major birth defects. Assisted hatching and assisted fertilization techniques have been implicated in increasing the incidence of monozygotic twinning.114, 116, 117 However, other studies attributed this problem to other aspects of ART, such as the gonadotropin treatment or the embryo culture conditions.118, 119, 120 Nevertheless, this phenomenon warrants further investigation because monozygotic twinning is more likely to result in miscarriage or other obstetrical complications.
Cytoplasm/germinal vesicle transfers
To remedy repeated implantation failures after ART treatments, another laboratory procedure has been described in which a small amount of cytoplasm is transferred from donor oocytes into the patients’ own oocytes.121, 122 The hypothesis underlying this procedure is that in some patients, certain defects in ooplasm may prevent the resultant embryo from normal development and implantation, and such defects may be corrected by infusion of cytoplasm from oocytes of fertile women. The importance of cytoplasmic factors in embryo development has been recognized for some time.123 Under certain culture conditions, embryos from outbred mice cannot develop beyond the two-cell stage (two-cell block). This block could be overcome after the embryos were injected with cytoplasm from inbred mouse embryos without the two-cell block. Mitochondria have been implicated as one of the candidates for the cytoplasmic regulator(s) of embryo development.124, 125, 126 Although successful pregnancies have resulted from cytoplasm transfers, there are no controlled studies to verify that it can improve the developmental potential of IVF-generated embryos. One study found this procedure ineffective in improving pregnancy rates in patients with diminished ovarian reserve.127 The caveat of this study, similar to many others evaluating the efficacy of ART, is that its study subjects were not randomized.
Based on the same hypothesis, an even bolder procedure has been proposed whereby the germinal vesicle from immature oocytes of infertile patients is removed and transplanted into enucleated oocyte from fertile donors.128, 129 To the best of our knowledge, no successful human pregnancy has been produced from this procedure. Both cytoplasm transfer and germinal vesicle transfer procedures can allow the patient’s genetic inheritance to be passed on to the offspring. However, clinical application of these techniques is not anticipated in the forseeable future because of ethical complications and safety concerns regarding these procedures.
MICROBIOPSY AND GENETIC DIAGNOSIS
The establishment of successful human pregnancies after preimplantation genetic diagnosis (PGD) was reported in 1990.130 Preimplantation genetic diagnosis became possible as a result of the coexistence of expertise in ART techniques and techniques for single-cell genetic analysis. PGD was initially developed to assist those carriers of severe genetic disorders who wish to have a healthy child without having to terminate an affected pregnancy. Indications for PGD continue to expand.131 By the year 2002, more than 5000 PGD cycles had been performed worldwide, resulting in the births of over 1000 infants. In the latest report of the European Society of Human Reproduction and Embryology PGD Consortium of 39 centers for the year 2005 alone, 3488 cycles of PGD were performed with 670 babies born. However, less than one third of these PGD cycles were performed for patients with genetic disorders and most of the cycles were performed to screen embryos for aneuloidy in an attempt to improve implantation rates.
born because of this new technology. The complete sequencing of the human genome and availability of more sensitive instruments will further accelerate technological advancement of PGD.132, 133 It can be anticipated that as public awareness of the genetic information grows, so will the demand for PGD.134, 135
Microbiopsy and embryologic work
Equipment and related facilities for biopsy are the same as for the use for ICSI and other embryo micromanipulative procedures. Methods for opening the ZP are the same as for assisted hatching, except that the opening may be slightly larger (approximately 30–40 μm). Strictly speaking, PGD can be performed either on oocytes prior to fertilization, also known as preconception diagnosis, or on embryos prior to the initiation of implantation. Procedures for polar body biopsy have been described by Verlinsky and colleagues.136 One of the advantages of preconception diagnosis is that polar bodies are not involved in subsequent embryo development and their removal, therefore, would cause little harm on embryonic development. Preconception diagnosis is advantageous also because the oocyte is not regarded as an individual and, therefore, manipulation on oocytes is more acceptable from religious or ethical points of view. However, this procedure is cumbersome because it often requires sequential biopsies of the first and second polar body to avoid misdiagnosis because of crossing-over.137 Furthermore, genetic disorders carried by the male partner cannot be detected by testing polar bodies of the oocyte.
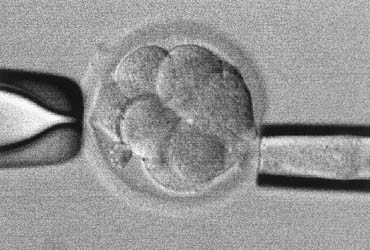
Biopsy of cleavage-stage embryos is probably the most common approach for PGD, wherein one or two cells (blastomeres) are removed from the embryo (Fig. 3). Blastomere biopsy should preferably be done no later than day 3 postfertilization because embryos start to compact around that time.138 The adhesion between blastomeres in a compacted embryo makes biopsy difficult. Compaction is mainly a function of cell-cell junctional proteins and is calcium/magnesium dependent.139, 140 By briefly exposing embryos to a calcium-magnesium-free medium, embryos can be decompacted, thus allowing the removal of one or two blastomeres without damaging the embryo.141 Decompaction can be reversed without harmful effect if the embryos are transferred back to the regular medium within 10 minutes. Embryo development is not significantly compromised by blastomere biopsy.142 Van de Velde and colleagues143 evaluated 188 PGD cycles and found that embryo survival was not different after removing one or two blastomeres from embryos with seven or more cells. The removal of two blastomeres for genetic analysis is preferred because it will help to produce a more accurate and reliable diagnosis.144 Frozen-thawed embryos, if they have survived well, can be used for biopsy. However, the combination of biopsy and cryopreservation can substantially reduce the embryo survival.145, 146 Therefore, “banking” embryos for future biopsy should be avoided.
|
In humans, the embryo usually reaches the blastocyst stage around day 5 postfertilization. Embryo culture media can now support the in vitro development of IVF-generated embryos to the blastocyst stage,147 making blastocyst biopsy feasible. Blastocyst biopsy has the potential to provide more cells for genetic analysis. It appears to cause less damage to the embryo.148, 149 The removal of trophoblast cells (the peripheral cells surrounding the blastocele cavity) may cause the blastocyst to collapse but the embryo can usually recover after the opening of blastocele cavity is resealed. Two factors limit the use of blastocyst biopsy for PGD. First, blastocyst biopsy can only be performed on day 5, which leaves little time for genetic analysis before the embryo must be transferred to the uterus. Second, mosaicism in trophectoderm is common, which will make diagnosis inaccurate.150 Blastocyst biopsy may become an option for PGD in the future when a fast and accurate genetic testing method emerges.
Chromosome analysis
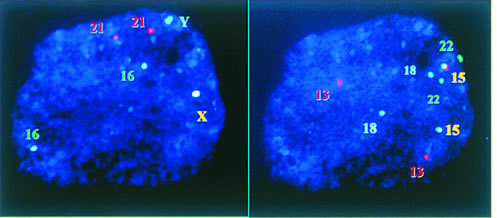
For most chromosomal analyses, the biopsied cell must be properly processed to remove as much cytoplasm as possible. One fixation method was first described by Tarkowsky151 and later modified by Griffin and colleagues152 and Munne and colleagues.153 In this method, the cell is swelled in a hypotonic solution and placed on a glass slide. The fixative (acetic acid and methanol) is then added drop by drop onto the cell. The impact of fixative drops helps to spread the cell and remove the cytoplasm. Another approach was reported by Coonen and colleagues141 and Harper and colleagues.154 In this method, the removal of cytoplasm is accomplished by exposing the cell to a hypotonic denaturing solution containing a detergent. Xu and coworkers155 compared the two methods, and found that both worked well with adequate training.155 More recently, Dozortsev and McGinnis156 suggested that a combination of the two methods works even better. Fixed nuclei are exposed to fluorescent-labeled DNA probes to allow the probes to hybridize with complimentary DNA sequences on the target chromosome, a process known as fluorescence in situ hybridization (FISH). Specific fluorescent signals can be visualized under a microscope with filters of appropriate wavelength. Up to nine chromosomes, for example, X, Y, 13, 14, 16, 18, 21, 22, can now be tested after two sequential FISH analyses in a single cell (Fig. 4).157, 158 Detection and elimination of chromosome aberrations in preimplantation-stage embryos first became possible in the early 1990s with the advent of the fast, sensitive, and accurate FISH technology152 and the availability of directly labeled probes. Indeed, FISH may be the easiest and most accurate way of determining the gender of an embryo, and it has the advantage of including additional DNA probes to screen other chromosomes.
|
Aneuploidy occurs in 10–30% of human conceptuses, mostly as a result of errors occurring during female meiosis. Aneuploidy accounts for at least one third of miscarriages and is the leading genetic cause of developmental disabilities and mental retardation.159 The frequency of aneuploidy in embryos increases with maternal age. An embryo’s morphologic appearance does not always reflect its genetic makeup because embryos with various kinds of aneuploidy and unbalanced chromosomes could well survive beyond the blastocyst stage in vitro.160, 161 Therefore, it seems desirable to screen for aneuploidy in preimplantation embryos so that only embryos with euploidy will be used to establish a pregnancy. Because aneuploidy is mainly attributed to female meiotic errors, screening polar bodies is a reasonable option.162, 163 The effectiveness of aneuploidy screening has been confounded with several difficulties. First, most candidates for aneuploidy screening often suffer diminished ovarian reserve because of advanced age. The decline in ovarian reserve begins in a woman in her late 20s.164 Consequently, these women do not respond well to ovarian stimulation, and often produce a limited number of oocytes and embryos for aneuploidy screening. Second, not all the cells in the early cleavage-stage embryo are carrying exact the same number of chromosomes. It is estimated that up to 40% of human embryos are mosaic,165 which will adversely affect the accuracy of PGD. Current aneuploidy screening methods also suffer from the limited number of chromosomes that can be examined on a single cell. Polymerase chain reaction (PCR)-based aneuploidy analyses, such as multiplex, fluorescent polymerase chain reaction (MF-PCR),166 and comparative genome hybridization (CGH)167, 168 can potentially screen all chromosomes in a single cell. However, their complexity and the lengthy time required to complete a test severely limit their routine use for PGD.169, 170
A retrospective, multicenter report indicates that preimplantation screening for aneuploidy can reduce the incidence of miscarriage but does not increase pregnancy rates after ART treatments.171 This finding was later confirmed in our randomized, controlled clinical trial.172 Taken together, screening for aneuploidy in preimplantation embryos may be beneficial under certain circumstances, but the effectiveness does not appear to justify its routine use for all patients. The decision for doing aneuploidy analysis, indeed, is a complex one. A risk-benefit evaluation should be performed to take into consideration multiple factors, such as the woman’s age, previous fertility history, response to stimulation, number and quality of embryos available for analysis, patient attitude toward miscarriages and abortion, etc. Perhaps, the single most important factor seems to be whether or not there is a selective advantage, that is, whether the number of embryos available for transfer exceeds the number of embryos acceptable for transfer without risking a higher-order multiple gestation. Aneuploidy screening may be recommended to those patients who have previous IVF failures with morphologically normal embryos, or those who have miscarriages because of known chromosomal anomalies.
In addition to aneuploidy, structural anomalies in the chromosome can also be detected by PGD. Structural chromosomal rearrangement may affect human reproduction. Schreurs and associates173 found that the prevalence of autosomal reciprocal balanced translocation was seven times higher in patients (1.14%) who cannot establish or maintain pregnancies than in the general population (0.16%). Prolonged culture of embryos to the blastocyst stage may be selective against embryos with balanced translocations because embryos carrying this type of translocations cannot develop normally in vitro.160, 174 However, blastocyst culture will not eliminate embryos with unbalanced translocations or other anomalies. First reports of PGD for chromosome translocation were published in the late 1990s.162, 175 Several strategies for chromosomal rearrangements have been attempted. Polar body biopsy and fluorescent whole-chromosome painting probes allow a direct assessment of chromosomal structure, thereby predicting the normalcy of the embryos. However, polar body chromosomes may easily clump together or degenerate, and thus not all polar bodies are informative. It is not an option if the male parent carries a chromosomal rearrangement. For the detection of chromosomal rearrangement, various types of DNA probes have been used. Breakpoint specific probes may provide the most informative diagnostic value,176 but these probes have to be made for each individual patient because they are carrier-specific. Therefore, it is less practical as the cost alone will prevent its wide application. Another more sophisticated approach is the so-called interphase-metaphase conversion, and its clinical application has been reported.177, 178 Telomeric probes are made from the DNA sequences located at the tip of each chromosome arm. Such probes have been used successfully for PGD.179, 180, 181 If the breakpoints are so close to the tip, for example, in the case of cryptic translocation, the only useful probes will be the telomeric ones. The commercial availability of telomeric probes for all human chromosomes has provided a convenient source of testing reagents. One of the limitations of telomeric probes is that they will not distinguish the balanced status from the normal ones.
DNA MUTATION ANALYSIS FOR MONOGENIC DISEASES
Once the precise diagnosis of a genetic disorder is made at the DNA level, it can be detected by PGD techniques. It is estimated that PGD attempts for more than 50 autosomal recessive conditions and autosomal dominant disorders have been reported from various PGD centers around the world, including cystic fibrosis,182 Huntington’s disease,183 sickle cell anemia,184 and early-onset Alzheimer disease,185 to name a few. Reports from two international working groups provide a glimpse of the activities in the area,134, 135 but they are quickly outdated. Knowledge of human genetic diseases has been accumulated astonishingly fast in the last several years as the Human Genome Project nears completion and this trend will certainly continue in the coming decade.132, 133
Compared to established prenatal diagnosis procedures, PGD has additional technical challenges because only a minute amount of DNA (approximately 6 pg of DNA per cell) is available for analysis. The advent of PCR technology made single-cell DNA analysis possible.186 Unlike chromosomal analysis, biopsied cells are simply placed individually into small test tubes for DNA analysis by PCR. Usually, a nested PCR (two sets of DNA primers and two rounds of PCR) is preferred in most centers because it will increase sensitivity. Although single-cell PCR usually is efficient, as high as 18% of the cells might not produce amplification signals.187 If multiple mutation sites are involved in a disorder, either a multiplex PCR or whole-genome amplification can be applied so that multiple loci can be analyzed simultaneously.188, 189, 190 One of the major challenges of this procedure is to prevent foreign DNA contamination, which can produce in a false result. Allele dropout (ADO) in PCR is another common cause for false results. In a situation of compound mutations, or when an autosomal dominant mutation is involved, ADO could lead to misdiagnosis with an affected pregnancy. In a very recent report on PCR for γ-globin gene, 8.6% of ADO was detected.191 To reduce the risk of ADO, linkage markers should be included in the PCR.144
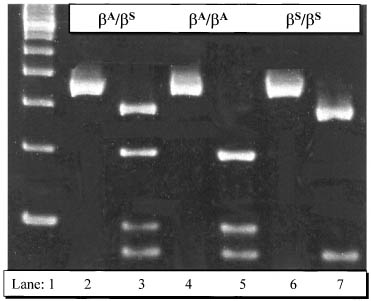
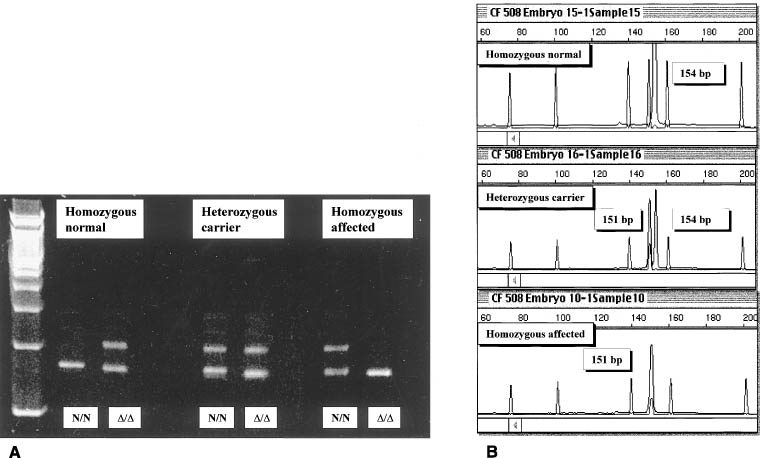
After DNA amplification with PCR, a classic and effective method to detect mutation is restriction enzyme analysis. Because many alterations in DNA sequence result in loss or gain of a restriction site, DNA from homozygous normal, heterozygous carrier or homozygous affected embryos can be easily distinguished after restriction digestion and electrophoresis of the PCR amplification product (Fig. 5). Alternatively, PCR primers can be designed so that embryonic DNA can be amplified selectively for or against the mutated sequence, and the presence or absence of the amplification product will indicate whether the sample is from an affected embryo. This approach is particularly useful for the detection of point mutations that do not affect a restriction site. A variety of other new modifications of PCR methods have been introduced. Fluorescent PCR is sensitive and has become popular. It can be used for detecting deletions (e.g., CF ΔF508 mutation) and insertions (for example TSD 1278ins4 mutation), replacing an old method called heteroduplex formation (Fig. 6).192 Fluorescent PCR also has been used for dealing with complex systems involving many loci.165 Real-time PCR refers to PCR systems that allow continuous measurement of the accumulation of a specific amplification product, and is useful in PGD because the electrophoretic analysis of PCR product is eliminated.193 Liquid chromatography can be used to sequence PCR product directly, which is particularly advantageous when the mutation does not involve a gain or loss of a restriction site.194
|
|
The list of indications for PGD has been grown rapidly131 and is no longer restricted to lethal genetic disorders. Mutations that predispose patients to certain diseases, such as cancer, have been screened for by PGD, even though a direct causal-effect relation has not been established between the mutations and the disease.195, 196 In one case, a healthy infant was born after a successful PGD for retinoblastoma (K. Xu and colleagues, April 2002, unpublished data). PGD for Alhzeimer’s disease is another indication that has been successfully carried out.185 Verlinsky and coworkers197 reported the first PGD case for avoiding Fanconi’s anemia, while at the same time selecting a HLA matching sibling donor for stem cell transplantation. This development is beneficial to many affected families, but at the same time has caused public debate. One cannot expect that there will be a uniform acceptance by the society for this type of new technology. However, it is certain that more couples will benefit from PGD in the future.
MICROMANIPULATION IN FUTURE ART
Micromanipulation techniques were originally developed to facilitate scientific investigations. Scientific breakthroughs have been made, and continue to be made with the aid of micromanipulation technologies. Some of the new technologies may lead to cures for many human diseases. However, whether they will have a place in future ART is dependent not only on further technical improvement but also on public acceptance. Medical professionals should be keenly aware of their professional as well as social and ethical responsibilities in the pursuit of technical advancement.
Cloning
Cloning is making a copy of an individual. The simplest approach is to split a cleavage-stage embryo and allow individual cells to grow into individual embryos. A sheep embryo was split and the two resulting halves of the embryo were transferred and gave rise to two identical sheep.198 Cloning can also be accomplished by transplanting the nucleus from a somatic cell into an enucleated oocyte, also known as somatic cell nuclear transfer (SCNT). The famous sheep, Dolly, was born as a result of SCNT.199 Cloning by SCNT offers great potential for scientific investigations of cell biology. It serves as an indispensable model for studying cytoplasm-nuclear interactions. Therapeutic cloning refers to SCNT technologies that can generate cells, tissues, or even organs to cure diseases. Using SCNT for reproductive purposes is not acceptable ethically by the majority of the public and is not safe, given the observations of many health problems in the cloned animals.200
Stem cell technology
Until a certain stage in development (day 3.5 in mice), embryonic cells are considered totipotent, that is, they have the potential to differentiate into any cell type found in the body. Such cells can be genetically engineered and used to generate tissues or organs with new genetic characteristics.201, 202 These cells can also be injected into a blastocyst and the resulting offspring could have cells or tissues derived from the genetically engineered stem cells. If the germ cells are derived from these cells, the unique genetic trait will become inheritable. Thus, the injection of stem cells with a specifically mutated gene can create a line of animals lacking the function of the gene, a technology called targeted mutagenesis or gene knockout.203 This technology has produced many animal models for studying various human diseases, ranging from cancer, to neurologic and immunologic disorders.204, 205, 206 Stem cells have been generated from human preimplantation embryos,207 paving the way for medical applications of the stem cell technologies. However, the generation of stem cells requires the destruction of live embryos, to which many in society strongly object on ethical or religious grounds.
Gene therapy
Foreign genes can be injected into the pronuclei of a newly fertilized embryo (zygote), and the injected genes can be incorporated into the embryo’s genome and be expressed.208 This technology is referred to as transgenesis. The generation of transgenic embryos or live animals provides a useful model system for studying an individual gene’s function and interaction with other genes. A specific trait of medical or economic value can be introduced to animals by transgenic technology. For example, a transgenic goat has been created to produce human plasminogen activator in its milk, which is a convenient source for isolating the therapeutic agent for treating stroke and heart attacks.209 However, the insertion of a foreign gene is poorly controlled, and as a result, the injected genes are not always functional or the insertion of the gene could interrupt the normal function of host genome. Until these problems are resolved, no attempt of transgenic technology should be made in humans.
Since the birth of the first IVF baby more than 30 years ago, ART has quickly become one of the most dynamic disciplines of medicine. Micromanipulation techniques have been, and will continue to be, an important catalyst for the rapid advancement in this area. Further development in this area will be dependent on improved understanding of human reproduction. More studies are also needed to evaluate the long-term benefits and risks of the current technologies.
ACKNOWLEDGMENT
We would like to thank Randall Barnes, MD, and Karen Horan, MSc, for their comments on the manuscript. We also wish to thank our respective colleagues at Northwestern University and Weil Cornell Medical College IVF programs for their assistance and support during the preparation of this manuscript.
REFERENCES
Chambers R, Chambers EL: Investigations into the Nature of Living Cell. Cambridge, MA, Harvard University Press, 1961 |
|
Slayter EM, Slayter HS (eds): Light and Electron Microscope. Cambridge, MA, Cambridge University, 1992 |
|
Lin TP: Egg micromanipulation. In Daniel JC (ed): Methods in Mammalian Embryology. pp 157, 171 San Francisco, Freeman and Company, 1971 |
|
Cohen J, Malter HE, Talansky BE, Grifo J (eds): Micromanipulation of Human Gametes and Embryos. New York, Raven Press, 1992 |
|
Van Steirteghem A, Joris H, Nagy P, et al: Assisted fertilization. In Ascosta AA, Kruger TF (eds): Human Spermatozoa in Assisted Reproduction. 2nd edition. pp 413, 432 New York, Parthenon Publishing, 1996 |
|
Smith SE, Tucker MJ: Micromanipulation for Assisted Reproduction. In Sciarra JJ (ed): Gynecology & Obstetrics. Vol 5:pp x, xx Chicago, Lippincott Williams & Wilkins, 1997 |
|
Burmeister L, Palermo GD, Rosenwaks Z: IVF: The new era. Int J Fertil Women Med 46:137, 2001 |
|
Gordon JW, Grunfeld L, Garrisi GJ, et al: Fertilization of human oocytes by sperm from infertile males after zonapellucida drilling. Fertil Steril 50:68, 1988 |
|
Malter HE, Cohen J: Partial zona dissection of the human oocyte: A nontraumatic method using micromanipulation to assist zona pellucida penetration. Fertil Steril 51:139, 1989 |
|
Strohmer H, Feichtinger W: Successful clinical application of laser for micromanipulation in an in vitro fertilization program. Fertil Steril 58:212, 1992 |
|
Fishel S, Antinori S, Jackson P, et al: Presentation of six pregnancies established by sub-zonal insemination (SUZI). Hum Reprod 6:124, 1991 |
|
Palermo G, Joris H, Devroey P, et al: Pregnancies after intracytoplasmic injection of single spermatozoon into an oocyte. Lancet 340:17, 1992 |
|
Uehara T, Yanagimachi R: Microsurgical injection of spermatozoa into hamster eggs with subsequent transformation of sperm nuclei into male pronuclei. Biol Reprod 15:467, 1976 |
|
Iritani A: Micromanipulation of gametes for in vitro assisted fertilization. Mol Reprod Devel 28:199, 1991 |
|
Ng SC, Bongso A, Ratnam SS: Microinjection of human oocytes: A technique for severe oligoasthenoteratozoospermia. Fertil Steril 56:1117, 1991 |
|
Yanagimachi R: Mammalian fertilization. In Knobil E, Neill JD (eds): The Physiology of Reproduction. Vol. 1:2nd ed. pp 189, 317 New York, Raven Press, 1994 |
|
Tesarik J, Sousa M, Testart J: Human oocyte activation after intracytoplasmic sperm injection. Hum Reprod 9:511, 1994 |
|
Tesarik J, Sousa M: Key elements of a highly efficient intracytoplasmic sperm injection technique: Ca2+ fluxes and oocyte cytoplasmic dislocation. Fertil Steril 64:770, 1995 |
|
Gearon CM, Taylor AS, Forman RG: Factors affecting activation and fertilization of human oocytes followingintracytoplasmic injection. Hum Reprod 10:896, 1995. |
|
Gerris J, Mangelschots K, Van Royen E, et al: ICSI and severe male-factor infertility: Breaking the sperm tail prior to injection. Hum Reprod 10:484, 1995. |
|
Dozortsev D, Rybouchkin A, De Sutter P, et al: Sperm plasma membrane damage prior to intracytoplasmic sperm injection: A necessary condition for sperm nucleus decondensation. Hum Reprod 10:2960, 1995 |
|
Levran D, Bider D, Yonesh M, et al: A randomized study of intracytoplasmic sperm injection (ICSI) versus subzonal insemination (SUZI) for the management of severe male-factor infertility. J Assist Reprod Genet 12:319, 1995 |
|
Tarlatzis BC, Bili H: Intracytoplasmic sperm injection. Survey of world results Ann NY Acad Sci 900:336, 2000 |
|
Kahraman S, Ozgur S, Alatas C, et al: High implantation and pregnancy rates with testicular sperm extraction and intracytoplasmic sperm injection in obstructive and non–obstructive azoospermia. Hum Reprod 11:673, 1996. |
|
Silber SJ, van Steirteghem A, Nagy Z, et al: Normal pregnancies resulting from testicular sperm extraction and intracytoplasmic sperm injection for azoospermia due to maturation arrest. Fertil Steril 66:110, 1996 |
|
Tournaye H, Liu J, Nagy Z, et al: The use of testicular sperm for intracytoplasmic sperm injection in patients with necrozoospermia. Fertil Steril 66:331, 1996 |
|
Devroey P, Silber S, Nagy Z, et al: Ongoing pregnancies and birth after intracytoplasmic sperm injection with frozen–thawed epididymal spermatozoa. Hum Reprod 10:903, 1995 |
|
Gil-Salom M, Romero J, Minguez Y, et al: Pregnancies after intracytoplasmic sperm injection with cryopreserved testicular spermatozoa. Hum Reprod 11:1309, 1996 |
|
Podsiadly BT, Woolcott RJ, Stanger JD, et al: Pregnancy resulting from intracytoplasmic injection of cryopreserved spermatozoa recovered from testicular biopsy. Hum Reprod 11:1306, 1996 |
|
Palermo GD, Schlegel PN, Colombero LT, et al: Aggressive sperm immobilization prior to intracytoplasmic sperm injection with immature spermatozoa improves fertilization and pregnancy rates. Hum Reprod 11:1023, 1996 |
|
Schoysman R, Vanderzwalmen P, Bertin G, et al: Oocyte insemination with spermatozoa precursors. Curr Opin Urol 9:541, 1999 |
|
Levran D, Nahum H, Farhi J, Weissman A: Poor outcome with round spermatid injection in azoospermic patients withmaturation arrest. Fertil Steril 74:443, 2000 |
|
Terriou P, Hans E, Giorgetti C et al: Pentoxifylline initiates motility in spontaneously immotile epididymal andtesticular spermatozoa and allows normal fertilization, pregnancy, and birthafter intracytoplasmic sperm injection. J Assist Reprod Genet. 2000 Apr;17(4):194-9. |
|
Kovacic B, Vlaisavljevic V, Reljic M: Clinical use of pentoxifylline for activation of immotile testicular sperm beforeICSI in patients with azoospermia. J Androl. 2006 Jan-Feb;27(1):45-52. |
|
Schroeder AC, Schultz RM, Kopf GS, et al: Fetuin inhibits zona pellucida hardening and conversion of ZP2 to ZP2f during spontaneous mouse oocyte maturation in vitro in the absence of serum. Biol Reprod 43:891, 1990 |
|
Zhang X, Rutledge J, Armstrong DT: Studies on zona hardening in rat oocytes that are matured in vitro in aserum-free medium. Mol Reprod Devel 28:292, 1991 |
|
Barnes FL, Crombie A, Gardner DK, et al: Blastocyst development and birth after in-vitro maturation of human primaryoocytes, intracytoplasmic sperm injection and assisted hatching. Hum Reprod 10:3243, 1995 |
|
Nagy ZP, Cecile J, Liu J, et al: Pregnancy and birth after intracytoplasmic sperm injection of in vitro matured germinal-vesicle stage oocytes: Case report. Fertil Steril 65:1047, 1996 |
|
Chian RC, Gulekli B, Buckett WM, Tan SL: Priming with human chorionic gonadotropin before retrieval of immature oocytes in women with infertility due to the polycystic ovary syndrome. N Engl J Med 341:1624, 1999 |
|
Vincent C, Pickering SJ, Johnson MH: The hardening effect of dimethylsulphoxide on the mouse zona pellucida requires the presence of an oocyte and is associated with a reduction in the number of cortical granules present. J Reprod Fertil 89:253, 1990 |
|
Manna C, Rienzi L, Greco E, et al: Zona pellucida solubility and cortical granule complements in human oocytes following assisted reproductive techniques. Zygote 9:201, 2001 |
|
Tucker MJ, Wright G, Morton PC, Massey JB: Birth after cryopreservation of immature oocytes with subsequent in vitro maturation. Fertil Steril 70:578, 1998 |
|
Kuleshova L, Gianaroli L, Magli C, et al: Birth following vitrification of a small number of human oocytes: Case report. Hum Reprod 14:3077, 1999 |
|
Wallace WH, Anderson RA, Irvine DS: Fertility preservation for young patients with cancer: who is at risk and what can be offered? Lancet Oncol. 2005 Apr;6(4):209-18. |
|
Rosendahl M, Loft A, Byskov AG et al: Biochemical pregnancy after fertilization of an oocyte aspirated from aheterotopic autotransplant of cryopreserved ovarian tissue: case report. Hum Reprod. 2006 Aug;21(8):2006-9. Epub 2006 May 9. |
|
Marina S, Marina F, Alcolea R, et al: Pregnancy following intracytoplasmic sperm injection from an HIV–1–seropositive man. Hum Reprod 13:3247, 1998 |
|
Loutradis D, Drakakis P, Kallianidis K, et al: Birth of two infants who were seronegative for human immunodeficiency virustype 1 (HIV-1) after intracytoplasmic injection of sperm from HIV–1–seropositive men. Fertil Steril 75:210, 2001 |
|
Chan AW, Luetjens CM, Dominko T, et al: Foreign DNA transmission by ICSI: injection of spermatozoa bound with exogenous DNA results in embryonic GFP expression and live rhesus monkey births. Mol Hum Reprod 6:26, 2000 |
|
Chan AW, Chong KY, Martinovich C, et al: Transgenic monkeys produced by retroviral gene transfer into mature oocytes. Science 291:309, 2001 |
|
Aboulghar MA, Mansour RT, et al: Prospective controlled randomized study of in vitro fertilization versus intracytoplasmic sperm injection in the treatment of tubal factor infertility with normal semen parameters. Fertil Steril 66:753, 1996 |
|
Gabrielsen A, Petersen K, Mikkelsen AL, et al: Intracytoplasmic sperm injection does not overcome an oocyte defect in previous fertilization failure with conventional in-vitro fertilization and normal spermatozoa. Hum Reprod 11:1963, 1996 |
|
Ola B, Afnan M, Sharif K, et al: Should ICSI be the treatment of choice for all cases of in-vitro conception? Considerations of fertilization and embryo development, cost effectiveness and safety Hum Reprod 16:2485, 2001 |
|
Najmabadi H, Huang V, Yen P, et al: Substantial prevalence of microdeletions of the Y–chromosome in infertile men with idiopathic azoospermia and oligospermia detected using a sequence tagged site based mapping strategy. J Clin Endcrinol Metab 81:1347, 1996 |
|
Pryor JL, Kent-First M, Muallem A, et al: Microdeletions in the Y chromosome of infertile men. N Engl J Med 336:534, 1997 |
|
Oliva R, Margarit E, Ballesc JP, et al: Prevalence of Y chromosome microdeletions in oligospermic and azoospermic candidates for intracytoplasmic sperm injection. Fertil Steril 70:506, 1998 |
|
Page DC, Silber S, Brown LG: Men with infertility caused by AZFc deletion can produce sons by intracytoplasmic sperm injection, but are likely to transmit the deletion and infertility. Hum Reprod 14:1722, 1999 |
|
Kent-First MG, Kol S, Muallem A, et al: The incidence and possible relevance of Y-linked microdeletions in babies born after intracytoplasmic sperm injection and their infertile fathers. Mol Hum Reprod 2:943, 1996 |
|
McInnes B, Rademaker A, Greene CA, et al: Abnormalities for chromosomes 13 and 21 detected in spermatozoa from infertile men. Hum Reprod 13:2787, 1998 |
|
Templado C, Hoang T, Greene C, et al: Aneuploid spermatozoa in infertile men: Teratozoospermia. Mol Reprod Dev 61:200, 2002 |
|
Macas E, Imthum B, Roselli M, et al: Chromosome analysis of single- and multipronucleated human zygotes proceeded after the intracytoplasmic injection procedure. J Assist Reprod Genet 13:345, 1996 |
|
Munné S, Márquez C, Reing A, et al: Chromosome abnormalities in embryos obtained after conventional in vitrofertilization and cytoplasmic sperm injection. Fertil Steril 69:904, 1998 |
|
Rosenbusch B, Sterzik K: Irregular chromosome segregation following ICSI? Hum Reprod 11:2337, 1996 |
|
Dumoulin JCM, Coonen E, Bras M, et al: Comparison of in-vitro development of embryos originating from either conventionalIVF or ICSI. Hum Reprod 15:402, 2000 |
|
Frattarelli JL, Leondires MP, Miller BT, et al: Intracytoplasmic sperm injection increases embryo fragmentation withoutaffecting clinical outcome. J Assist Reprod Genet 17:207, 2000 |
|
Griffiths TA, Murdoch AP, Herbert M: Embryonic development in vitro is compromised by the ICSI procedure. Hum Reprod 15:1592, 2000 |
|
Moilanen JM, Tulppala M, Rema I, et al: Fertilization, embryo quality, and cryosurvival in in vitro fertilizationand intracytoplasmic sperm injection cycles. J Assist Reprod Genet 16:17, 1999 |
|
Westphal LM, Hinckley MD, Behr B, et al: ICSI does not compromise blastocyst development. Fertil Steril 77:S14, 2002 |
|
Plachot M, Belaisch-Allart J, Mayenga JM, et al: Outcome of conventional IVF and ICSI on sibling oocytes in mild male factorinfertility. Hum Reprod 17:362, 2002 |
|
Nagy ZP, Liu J, Joris H, et al: The influence of the site of sperm deposition and mode of oolemma breakage at intracytoplasmic sperm injection on fertilization and embryo development rates. Hum Reprod 10:3171, 1995 |
|
Dumoulin JM, Coonen E, Bras M, et al: Embryo development and chromosomal anomalies after ICSI: Effect of the injection procedure. Hum Reprod 16:306, 2001 |
|
Hewitson L, Dominko T, Takahashi D, et al: Unique checkpoints during the first cell cycle of fertilization after intracytoplasmic sperm injection in rhesus monkeys. Nat Med 5:431, 1999 |
|
Hardarson T, Lundin K, Hamberger L: The position of the metaphase II spindle cannot be predicted by the location of the first polar body in the human oocyte. Hum Reprod 15:1372, 2000 |
|
Wang WH, Meng L, Hackett RJ, et al: The spindle observation and its relationship with fertilization after intracytoplasmic sperm injection in living human oocytes. Fertil Steril 75:348, 2001 |
|
Barak Y, Menezo Y, Veiga A, Elder K: A physiological replacement for polyvinylpyrrolidone (PVP) in assisted reproductive technology. Hum Fertil 4:99, 2001 |
|
Palermo GD, Neri QV, Hariprashad JJ, et al: ICSI and its outcome. Semin Reprod Med 18:161, 2000 |
|
Foresta C, Ferlin A: Offspring conceived by intracytoplasmic sperm injection. Lancet 358:1270, 2001 |
|
Bonduelle M, Liebaers I, Deketelaere V, et al: Neonatal data on a cohort of 2889 infants born after ICSI (1991–1999) and of 2995infants born after IVF (1983–1999). Hum Reprod 17:671, 2002 |
|
Van Steirteghem A, Bonduelle M, Liebaers I, et al: Children born after assisted reproductive technology. Am J Perinatol 19:59, 2002 |
|
Hansen M, Kurinczuk JJ, Bower C, et al: The risk of major birth defects after intracytoplasmic sperm injection and in vitro fertilization. N Engl J Med 346:725, 2002 |
|
Bowen JR, Gibson FL, Leslie GI, et al: Medical and developmental outcomes at 1 year for children conceived by intracytoplasmic sperm injection. Lancet 315:1529, 1998 |
|
Bonduelle M, Joris H, Hofmans K, et al: Mental development of 201 ICSI children at 2 years of age. Lancet 22:1553, 1998 |
|
Cox GF, Burger J, Lip V et al: Intracytoplasmic sperm injection may increase the risk of imprinting defects. Am J Hum Genet. 2002 Jul;71(1):162-4. |
|
Ludwig M, Katalinic A, Gross S et al: Increased prevalence of imprinting defects in patients with Angelman syndromeborn to subfertile couples. J Med Genet. 2005 Apr;42(4):289-91. |
|
Neri QV, Takeuchi T, Palermo GD: An update of assisted reproductive technologies results in the United States. Ann N Y Acad Sci. 2008 Apr;1127:41-8. |
|
Kanber D, Buiting K, Zeschnigk M et al: Low frequency of imprinting defects in ICSI children born small for gestationalage. Eur J Hum Genet. 2009 Jan;17(1):22-9. Epub 2008 Oct 22. |
|
Cohen J, Elsner C, Kort H, et al: Impairment of the hatching process following IVF in the human and improvement of implantation by assisted hatching using micromanipulation. Hum Reprod 5:7, 1990 |
|
Cohen J: Assisted hatching of human embryos. J IVF-ET 8:179, 1991 |
|
Orsini MW: Factors affecting loss of the zona pellucida in the hamster. J Anat 99:922, 1965 |
|
Orsini MW, Mclaren A: Loss of the zona pellucida in mice, and the effect tubal ligation and ovariectomy. J Reprod Fertil 13:485, 1967 |
|
Confino E, Rawlins R, Binor Z, Radwanska E: The effect of the oviduct, uterine, and in vitro environments on zona thinning in the mouse embryo. Fertil Steril 68:164, 1997 |
|
McDougall K, Hedrick JL, Bavister BD: In situ pH measurements of the Syrian hamster uterus during early pregnancy to determine the role of pH in zona pellucida loss in vivo. Reprod Fertil Develop 12:105, 2000 |
|
Gonzales DS, Bavister BD: In utero and in vitro proteinase activity during the Mesocricetus auratus embryo zona escape time window. Biol Reprod 64:222, 2001 |
|
Cannon MJ, Menino AR Jr: Changes in the bovine zona pellucida induced by plasmin or embryonic plasminogen activator. Mol Reprod Dev 51:330, 1998 |
|
Loret De Mola JR, Garside WT, et al: Analysis of the human zona pellucida during culture: correlation with diagnosis and the preovulatory hormonal environment. J Assist Reprod Genet 14:332, 1997 |
|
Garside WT, Loret de Mola JR, Bucci JA, et al: Sequential analysis of zona thickness during in vitro culture of human zygotes: correlation with embryo quality, age, and implantation. Mol Reprod Dev 47:99, 1997 |
|
Palmstierna M, Murkes D, Csemiczky G, et al: Zona pellucida thickness variation and occurrence of visible mononucleated blastomers in preembryos are associated with a high pregnancy rate in IVF treatment. J Assist Reprod Genet 15:70, 1998 |
|
Gabrielsen A, Lindenberg S, Petersen K: The impact of the zona pellucida thickness variation of human embryos on pregnancy outcome in relation to suboptimal embryo development. A prospective randomized controlled study Hum Reprod 16:2166, 2001 |
|
Cieslak J, Ivakhnenko V, Wolf G, et al: Three-dimensional partial zona dissection for preimplantation genetic diagnosis and assisted hatching. Fertil Steril 71:308, 1999 |
|
Khalifa EA, Tucker MJ, Hunt P: Cruciate thinning of the zona pellucida for more successful enhancement of blastocyst hatching in the mouse. Human Reprod 7:532, 1992 |
|
Blake DA, Forsberg AS, Johansson BR, et al: Laser zona pellucida thinning: An alternative approach to assisted hatching. Hum Reprod 16:1959, 2001 |
|
Tucker MJ, Luecke NM, Wiker SR, et al: Chemical removal of the outside of the zona pellucida of day 3 human embryos has no impact on implantation rate. J Assist Reprod Genet 10:187, 1993 |
|
Fong CY, Bongso A., Ng SC, et al: Ongoing normal pregnancy after transfer of zona-free blastocysts: Implications for embryo transfer in the human. Hum Reprod 12:557, 1997 |
|
Balaban B, Urman B, Alatas C, et al: A comparison of four different techniques of assisted hatching. Hum Reprod 17:1239, 2002 |
|
Cohen J, Alikani M, Trowbridge J, Rosenwaks Z: Implantation enhancement by selective assisted hatching using zona drilling of human embryos with poor prognosis. Hum Reprod 7:685, 1992 |
|
Schoolcraft WB, Schlenker T, Jones GS, et al: In vitro fertilization in women age 40 and older: the impact of assisted hatching. J Assist Reprod Genet 12:581, 1995 |
|
Stein A, Rufas O, Amit S, et al: Assisted hatching by partial zona dissection of human pre-embryos in patientswith recurrent implantation failure after in vitro fertilization. Fertil Steril 63:838, 1995 |
|
Antinori S, Selman HA, Caffa B, et al: Zona opening of human embryos using a non-contact UV laser for assisted hatching in patients with poor prognosis of pregnancy. Hum Reprod 11:2488, 1996 |
|
Meldrum DR, Wisot A, Yee B, et al: Assisted hatching reduces the age-related decline in IVF outcome in women younger than age 43 without increasing miscarriage or monozygotic twinning. J Assist Reprod Genet 15:418, 1998 |
|
Hellebaut S, De Sutter P, Dozortsev D, et al: Does assisted hatching improve implantation rates after in vitro fertilization or intracytoplasmic sperm injection in all patients? A prospective randomized study J Assist Reprod Genet 13:19, 1996 |
|
Lanzendorf SE, Nehchiri F, Mayer JF, et al: A prospective, randomized, double-blind study for the evaluation of assisted hatching in patients with advanced maternal age. Hum Reprod 13:409, 1998 |
|
Hurst B, Tucker M, Awoniyi A, Schlaff W: Assisted hatching does not enhance IVF success in good prognosis patients. J Assist Reprod Genet 15:62, 1998 |
|
Edirisinghe WR, Ahnonkitpanit V, Promviengchai S, et al: A study failing to determine significant benefits from assisted hatching: Patients selected for advanced age, zonal thickness of embryos, and previous failed attempts. J Assist Reprod Genet 16:294, 1999 |
|
American Society for Reproductive Medicine: The role of assisted hatching in IVF: A review of the literature (A Practice Committee Report). 2000 |
|
Slotnick RN, Ortega JE: Monoamniotic twinning and zona manipulation: A survey of U.S. IVF centerscorrelating zona manipulation procedures and high-risk twinning frequency J Assist Reprod Genet 13:381, 1996 |
|
Hershlag A, Paine T, Cooper GW, et al: Monozygotic twinning associated with mechanical assisted hatching. Fertil Steril 71:144, 1999 |
|
Sills ES, Moomjy M, Zaninovic N, et al: Human zona pellucida micromanipulation and monozygotic twinning frequency after IVF. Hum Reprod 15:890, 2000 |
|
Sheen TC, Chen SR, Au HK, et al: Herniated blastomere following chemically assisted hatching may result inmonozygotic twins. Fertil Steril 75:442, 2001 |
|
Schieve LA, Meikle SF, Peterson HB, et al: Does assisted hatching pose a risk for monozygotic twinning in pregnancies conceived through in vitro fertilization? Fertil Steril 74:288, 2000 |
|
Behr B, Fisch JD, Racowsky C, et al: Blastocyst-ET and monozygotic twinning. J Assist Reprod Genet 17:349, 2000 |
|
Schachter M, Raziel A, Friedler S, et al: Monozygotic twinning after assisted reproductive techniques: A phenomenon independent of micromanipulation. Hum Reprod 16:1264, 2001 |
|
Cohen J, Scott R, Schimmel T, et al: Birth of infant after transfer of anucleate donor oocyte cytoplasm into recipient eggs. Lancet 350:186, 1997 |
|
Lanzendorf SE, Mayer JF, Toner J, et al: Pregnancy following transfer of ooplasm from cryopreserved-thawed donoroocytes into recipient oocytes. Fertil Steril 71:575, 1999 |
|
Muggleton-Harris A, Whittingham DG, Wilson L: Cytoplasmic control of preimplantation development in vitro in the mouse. Nature 299:460, 1982 |
|
Brenner CA, Wolny YM, Barritt JA: Mitochondrial DNA deletion in human oocytes and embryos. Mol Hum Reprod 4:887, 1998 |
|
Van Blerkom J, Sinclair J, Davis P: Mitochondrial transfer between oocytes: Potential applications of mitochondrial donation and the issue of hteroplasmy. Hum Reprod 13:2857, 1998 |
|
Bavister BD, Squirrell JM: Mitochondrial distribution and function in oocytes and early embryos. Hum Reprod 15:189, 2000 |
|
Opsahl MS, Thorsell LP, Geltinger ME, et al: Donor oocyte cytoplasmic transfer did not enhance implantation of embryos of women with poor ovarian reserve. J Assist Reprod Genet 19:113, 2002 |
|
Krey L, Liu H, Zhang J, Grifo J: Fertility and maternal age strategies to improve pregnancy outcome. Ann NY Acad Sci 943:26, 2001 |
|
Moffa F, Comoglio F, Krey LC, et al: Germinal vesicle transfer between fresh and cryopreserved immature mouse oocytes. Hum Reprod 17:178, 2002 |
|
Handyside AH, Kontogianni EH, Hardy K, et al: Pregnancies from biopsied human preimplantation embryos sexed by Y-specific DNA amplification. Nature 344:768, 1990 |
|
Simpson JL: Changing indications for preimplantation genetic diagnosis (PGD). Mol Cell Endocrinol 183:69, 2001 |
|
International Human Genome Sequencing Consortium: Initial sequencing and analysis of the human genome. Nature 409:860, 2001 |
|
Venter JC, Adams MD, Myers J, et al: The sequence of the human genome. Science 291:1304, 2001 |
|
European Society for Human Reproduction and Embryology-Preimplantation Genetic Diagnosis Consortium: Data collection III (May 2001). Hum Reprod 17:233, 2002 |
|
International Working Group on Preimplantation Genetics:Preimplantation diagnosis: Experience of three thousand clinical cycles. Reprod Bio Med Online 3:49, 2001 |
|
Verlinsky Y, Ginsberg N, Lifchez A, et al: Analysis of the first polar body: Preconception genetic diagnosis. Hum Reprod 5:826, 1990 |
|
Angell RR: Polar body analysis: Possible pitfalls in preimplantation diagnosis of chromosomal disorders based on polar body analysis. Hum Reprod 9:181, 1994 |
|
Nikas G, Ao A, Winston RML, et al: Compaction and suerface polarity in the human embryo in vitro. Biol Reprod 55:32, 1995 |
|
Dale B, Gualtieri R, Talevi R, et al: Intercellular communication in the early human embryo. Mol Reprod Dev 29:22, 1991 |
|
Minami N, Takahashi A, Yamada M, et al: E-Cadherin localization and gap junction assembly in the mouse embryos are accelerated by the oviduct in vitro. Biol Reprod 54:170, 1996 |
|
Coonen E, Dumoulin JCM, Ramaekers FCS, et al: Optimal preparation of preimplantation embryo interphase nuclei for analysis by fluorescence in-situ hybridization. Hum Reprod 9:533, 1994 |
|
Hardy K, Martin KL, Leese HJ, et al: Human preimplantation development in vitro is not adversely affected by biopsy at the 8-cell stage. Hum Reprod 5:708, 1990 |
|
Van de Velde H, De Vos A, Sermon K, et al: Embryo implantation after biopsy of one or two cells from cleavage-stage embryos with a view to preimplantation genetic diagnosis. Prenat Diagn 20:1030, 2000 |
|
Lewis CM, Pinel T, Whittaker JC, et al: Controlling misdiagnosis errors in preimplantation genetic diagnosis: A comprehensive modelencompassing extrinsic and intrinsic sources of error. Hum Reprod 16:43, 2001 |
|
Joris H, Van den Abbeel E, et al: Reduced survival after human embryo biopsy and subsequent cryopreservation. Hum Reprod 14:2833, 1999 |
|
Magli MC, Gianaroli L, Fortini D, et al: Impact of blastomere biopsy and cryopreservation techniques on human embryo viability. Hum Reprod 14:770, 1999 |
|
Gardner DK: Development of serum-free media for the culture and transfer of human blastocysts. Hum Reprod 13:218, 1998 |
|
Dokras A, Sargent IL, Ross C, et al: Human trophectoderm biopsy and secretion of chorionic gonadotrophin. Hum Reprod 6:1453, 1991 |
|
Carson SA, Gentry WL, Smith AL: Trophectoderm microbiopsy in murine blastocysts: Comparison of four methods. J Assist Reprod Genet 10:427, 1993 |
|
Eviskov S, Verlinskt Y: Mosaicism in the inner cell mass of human blastocysts. Hum Reprod 13:3151, 1998 |
|
Tarkowsky AK: An air drying method for chromosome preparation from mouse eggs. Cytogenetics 5:394, 1964 |
|
Griffin DK, Handyside AH, Penketh RJA, et al: Fluorescent in-situ hybidization to interphase nuclei of human preimplantation embryos with X and Y chromosome specific probes. Hum Reprod 6:101, 1991 |
|
Munne S, Weier HU, Stein J, et al: A fast and efficient method for simultaneous X and Y in situ hybridization ofhuman blastomeres. J Assist Reprod Genet 10:82, 1993 |
|
Harper JC, Coonen E, Ramaekers FCS, et al: Identification of the sex of human preimplantation embryos in two hours using an improved spreading method and fluorescent in-situ hybridization (FISH) using directly labeled probes. Hum Reprod 9:721, 1994 |
|
Xu KP, Huang TH, Liu TZ, et al: Improving the fixation method for preimplantation genetic diagnosis by fluorescent insitu hybridization. J Assist Reprod Genet 15:570, 1998 |
|
Dozortsev DI, McGinnis KT: An improved fixation technique for fluorescent in situ hybridization for preimplantation genetic diagnosis. Fertil Steril 76:186, 2001 |
|
Munne S, Magli C, Bache M, et al: Preimplantation diagnosis of the aneuploidies most commonly found in spontaneous abortions and live births: XY,13,14,15,16,18,21,22. Prenat Diagn 18:1459, 1998 |
|
Gianaroli L, Magli MC, Ferraretti AP, et al: Preimplantation diagnosis for aneuploidies in patients undergoing in vitro fertilization with a poor prognosis: Identification of the categories for which it should be proposed. Fertil Steril 72:837, 1999 |
|
Hassold T, Hunt P: To err (meiotically) is human: The genesis of human aneuploidy. Nat Rev Genet 2:280, 2001 |
|
Menezo YJ, Bellec V, Zarouckian A, et al: Embryo selection by IVF, co-culture and transfer at the blastocyst stage in case of translocation. Hum Reprod 12:2802, 1997 |
|
Magli MC, Gianaroli L, Ferraretti AP: Chromosomal abnormalities in embryos. Mol Cell Endocrinol 183:S29, 2001 |
|
Durban M, Benet J, Egozcue J, et al: Chromosome studies in first polar bodies from hamster and human oocytes. Hum Reprod 13:583, 1998 |
|
Verlinsky Y, Cieslak J, Ivakhnenko V, et al: Chromosomal abnormalities in the first and second polar bodies. Mol Cell Endocrinol 183:S47, 2001 |
|
Dunson DB, Colombo B, Baird DD: Changes with age in the level and duration of fertility in the menstrual cycle. Hum Reprod 17:1399, 2002 |
|
Bielanska M, Tan SL, Ao A: Chromosomal mosaicism throughout human preimplantation development in vitro: Incidence, type, and relevance to embryo outcome. Hum Reprod 17:413, 2002 |
|
Findlay I, Matthews PL, Mulcahy BK, et al: Using MF-PCR to diagnose multiplee defects from single cells: Implications for PGD. Mol Cell Endocrinol 183:S5, 2001 |
|
Voullaire L, Slater H, Williamson R, et al: Chromosome analysis of blastomeres from human embryos by using comparative genomic hybridization. Hum Genet 106:210, 2000 |
|
Wells D, Delhanty JDA: Comprehensive chromosomal analysis of human preimplantation embryos using whole genome amplification and single cell comparative genomic hybridization. Mol Hum Reprod 6:1055, 2000 |
|
Weier HUG, Munne S, Lersch RA, et al: Towards a full karyotype screening of interphase cells: ‘FISH and chip’ technology. Mol Cell Endocrinol 183:S41, 2001 |
|
Malmgren H, Sahlen S, Inzunza J, et al: Single cell CGH analysis reveals a high degree of mosaicism in human embryos from patients with balanced structural chromosome aberrations. Mol Hum Reprod 8:502, 2000 |
|
Munne S, Magli C, Cohen J: Positive outcome after preimplantation diagnosis of aneuploidy in human embryos. Hum Reprod 14:2191-9, 1999 |
|
Milad MP, Zhang XJ, Pergament E, et al: Impact of preimplantation genetic diagnosis (PGD) screening for common aneuploidies on IVF pregnancy rates. Fertil Steril 76:P70, 2001 |
|
Schreurs A, Legius E, Meuleman C, et al: Increased frequency of chromosomal abnormalities in female partners of couples undergoing in vitro fertilization or intracytoplasmic sperm injection. Fertil Steril 74:94, 2000 |
|
Iwarsson E, Malmgren H, Inzunza J: Highly abnormal cleavage divisions in preimplantation embryos from translocation carriers. Prenat Diagn 20:1038, 2000 |
|
Munne S, Scott R, Sable D, Cohen J: First pregnancies after preconception diagnosis of translocations of maternal origin. Fertil Steril 69:675, 1998 |
|
Cassel MJ, Munne S, Fung J, et al: Carrier–specific breakpoint-spanning DNA probes: an approach to preimplantatin genetic diagnosis in interphase cells. Hum Reprod 12:2019, 1997 |
|
Evsikov S, Verlinsky Y: Visualization of chromosome in single human blastomeres. J Assist Reprod Genet 16:133, 1999 |
|
Willadsen SM, Levron J, Munne S, et al: Rapid visualization of metaphase chromosomes in single human blastomeres after fusion with in-vitro matured bovine eggs. Hum Reprod 14:470, 1999 |
|
Ning Y, Roschke A, Smith ACM, et al: A complete set of human telomeric probes and their clinical application. National Institute of Health and Institue of Molecular Medicine collaboration Nat Genet 14:486, 1996 |
|
Xu KP, Rosenwaks Z: Preimplantation genetic diagnosis (PGD): Challenges and potentials. In New MI (ed): Diagnosis and Treatment of the Unborn Child. pp 109, 116 New York, Idelson-Gnocchi Ltd. Publishers, 1999 |
|
Coonen E, Martini E, Dumoulin JC, et al: Preimplantation genetic diagnosis of a reciprocal translocation t(3;11)(q27.3;q24.3) insiblings Mol Hum Reprod 6:199, 2000 |
|
Handyside AH, Lesko JG, Tar’in JJ, et al: Birth of a normal girl after in vitro fertilization and preimplantation diagnostic testing for cystic fibrosis. N Engl J Med 327:905, 1992 |
|
Sermon K, Goossens V, Seneca S, et al: Preimplantation diagnosis for Huntington’s disease (HD): Clinical application and analysis of the HD expansion in affected embryos. Prenat Diagn 18:1427, 1998 |
|
Xu KP, Shi ZM, Veeck LL, et al: First unaffected pregnancy using preimplantation genetic diagnosis for sickle cell anemia. JAMA 281:1701, 1999 |
|
Verlinsky Y, Rechitsky S, Verlinsky O, et al: Preimplantation diagnosis for early-onset Alzheimer disease caused by V717L mutation. JAMA 287:1018, 2002 |
|
Mullis K B, Faloona F: Specific synthesis of DNA in vitro via a polymerase catalyzed chain reaction. Methods Enzymol 155:335, 1987 |
|
European Society for Human Reproduction and Embryology-Preimplantation Genetic Diagnosis consortium: Preliminary assessment of data from January 1997 to September 1998. Hum Reprod 14:3138, 1999 |
|
Zhang L, Cui X, Schmitt K, et al: Whole genome amplification from a single cell: Implications for genetic analysis. Proc Natl Acad Sci USA 89:5847, 1992 |
|
Xu KP, Tang YX, Grifo JA, et al: Primer extension preamplification for detection of multiple genetic loci from single human blastomeres. Hum Reprod 8:2206, 1993 |
|
Kristjansson K, Chong SS, Van den Veyver IB, et al: Preimplantation single cell analyses of dystrophin gene deletions using whole genome amplification. Nat Genet 6:19, 1994 |
|
Chamayou S, Alecci C, Ragolia C, et al: Successful application of preimplantation genetic diagnosis for β-thalassemia and sickle cell anaemia in Italy. Hum Reprod 17:1158, 2002 |
|
Ao A, Ray P., Harper J, et al: Clinical experience with preimplantation genetic diagnosis of cystic fibrosis (delta F508). Prenat Diag 16:137, 1996 |
|
Pierce KE, Rice JE, Sanchez JA, et al: Real-time OCR using molecular beacons for accurate detection of the Y chromosome in single human blastomere. Mol Hum Reprod 6:1155, 2000 |
|
Drury KC, Liu MC, Lilleberb S, et al: Results on single cell PCR for Huntington’s gene and WAVE(TM) product analysis for preimplantation genetic diagnosis. Mol Cell Endocrinol 183:S1, 2001 |
|
Ao A, Wells D, Handyside A, et al: Preimplantation genetic diagnosis of inherited cancer: Familial adenomatous polyposis coli. J Assist Reprod Genet 15:140, 1998 |
|
Verlinsky Y, Rechitsky S, Verlinsky O, et al: Preimplantation diagnosis for p53 tumour suppressor gene mutations. Reprod Bio Med Online 2:102, 2001 |
|
Verlinsky Y, Rechitsky S., Schoolcraft W, et al: Preimplantation diagnosis for Fanconi anemia combined with HLA matching. JAMA 285:3130, 2001 |
|
Willadsen SM: A method for culture of micromanipulated sheep embryos and its use to produce monozygotic twins. Nature 277:298, 1979 |
|
Wilmut I, Schnieke AE, McWhir J, et al: Viable offspring derived from fetal and adult mammalian cells. Nature 385:810, 1997 |
|
Wilmut I: Are there any normal cloned mammals? Nat Med 8:215, 2002 |
|
Rossant J: Stem cells from the mammalian blastocyst. Stem Cell 19:477, 2001 |
|
Rossant J, Bernelot-Moens C, Nagy A: Genome manipulation in embryonic stem cells. Phil Trans R Soc London B: Biol Sci 339:207, 1993 |
|
Robertson EJ: Using embryonic stem cells to introduce mutations into the mouse germ line. Biol Reprod 44:238, 1991 |
|
Janus C, Phinney AL, Chishti MA, Westaway D: New developments in animal models of Alzheimer’s disease. Curr Neurol Neurosci Rep 1:451, 2001 |
|
Deng CX, Brodie SG: Knockout mouse models and mammary tumorigenesis. Semin Cancer Biol 11:387, 2001 |
|
Mak T W, Penninger J M, Ohashi P S: Knockout mice: A paradigm shift in modern immunology. Nat Rev Immunol 1:11, 2001 |
|
Thomson JA, Itskovitz-Eldor J, Shapiro SS, et al: Embryonic stem cell lines derived from human blastocysts. Science 282:1145, 1998 |
|
Palmiter RD, Norstedt G, Gelinas RE, et al: Metallothionein-human GH fusion genes stimulate growth of mice. Science 222:809, 1983 |
|
Ebert KM, Selgrath JP, DiTullio P, et al: Transgenic production of a variant of human tissue-type plasminogen activator in goat milk: Generation of transgenic goats and analysis of expression. Biotechnology 9:835, 1991 |