The Posterior Pituitary Pathway
Authors
INTRODUCTION
In 1838, Rathke described the human hypophysis cerebri as being derived from two parts: an ectodermal dorsal invagination of the oral epithelium, which becomes the adenohypophysis, and a ventral process arising from the floor of the diencephalon, constituting the neurohypophysis or posterior pituitary gland (Figure 1). The vasopressor and oxytocic activities of the posterior pituitary were first shown by Oliver and Schafer in 1895 and by Dale in 1906. Subsequently, posterior pituitary extracts were shown to have antidiuretic and galactokinetic properties. Thereafter, these properties were reported to be due to two separate components, which eventually led to the structural analysis and synthesis of the two active peptide hormones and their binding proteins, the neurophysins, all of which are now clearly established to be secreted by the hypothalamoneurohypophysial system.
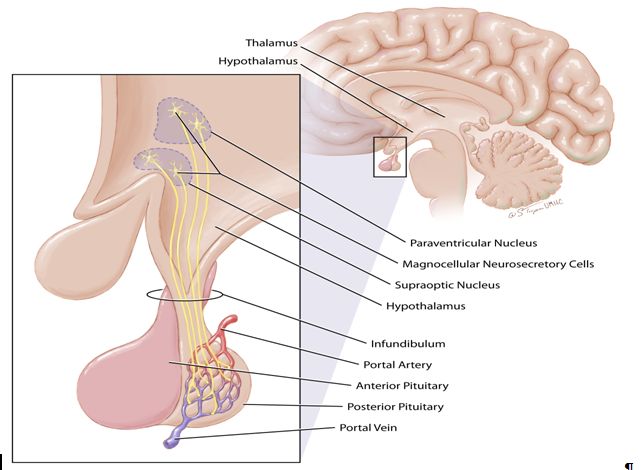
Fig. 1. The posterior pituitary. The posterior pituitary lobe or neurohypophysis arises as an extension of the floor of the diencephalon (thalamus and hypothalamus) and has direct continuity with the nuclei of the hypothalamus. The cell bodies of the magnocellular neurons extend through the infundibulum to discharge their contents into the portal circulation of the pituitary
STRUCTURE OF PITUITARY PEPTIDES
The structures of oxytocin and vasopressin are shown in Figure 2. Both oxytocin and vasopressin consist of nine amino acid residues, of which two are half cystines forming a disulfide bridge between positions 1 and 6. The human and other mammals have arginine vasopressin and not lysine vasopressin which has so far only been found in pigs (Figure 1). In the animal kingdom, there are six posterior pituitary hormones with oxytocic properties: oxytocin, mesotocin, isotocin, glumitocin, valitocin, and aspargtocin. Substitutions have occurred only in positions 3, 4, and 8, which suggests that the amino acid residues in positions 1, 2, 5, 6, 7, and 9 are essential for oxytocin function and could be obligatory components of the original ancestral molecule that has evolved to become oxytocin in mammals. The molecular weights of oxytocin and arginine vasopressin are 1007 and 1084 kDa, respectively.
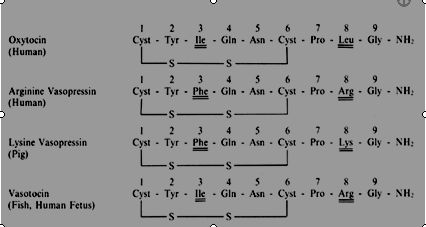 Fig. 2. Amino acid sequence of oxytocin, arginine vasopressin, lysine vasopressin, and vasotocin. Vasotocin is found in the human fetus. Lysine vasopressin is found only in pigs
Fig. 2. Amino acid sequence of oxytocin, arginine vasopressin, lysine vasopressin, and vasotocin. Vasotocin is found in the human fetus. Lysine vasopressin is found only in pigs
The neurophysins are polypeptides with a molecular weight of about 10,000 kDa. In the human, two forms of circulating neurophysins exist: estrogen-stimulated neurophysin (neurophysin I) and nicotine-stimulated neurophysin (neurophysin II).
BIOSYNTHESIS OF NEUROHYPOPHYSIAL PEPTIDES
The posterior pituitary peptides are synthesized primarily in the magnocellular neurons of the paraventricular and supraoptic nuclei of the hypothalamus. Although the paraventricular nucleus has been thought to be primarily responsible for oxytocin synthesis, while the supraoptic nucleus was responsible for vasopressin synthesis, there is overlap between them.1 Vasopressin and oxytocin are synthesized within separate neurosecretory cells, specialized to synthesize only that particular neurohypophysial hormone together with its neurophysin, although these separate neurons are found within each of these hypothalamic nuclei.2 The oxytocinergic and vasopressinergic neurons within the paraventricular and supraoptic nuclei are partially segregated within each nucleus, with oxytocinergic cells clustering rostrally, while vasopressin-containing cells are found more caudally.3
The oxytocin and vasopressin genes for humans and several other species have been isolated and determined.4 All have similar structural organization, confirming a common origin for both peptides from a mutual ancestral gene that separated into two genes for the two hormones about 400 million years ago when vertebrates evolved.5 In all species (mouse, rat, cow, and human) so far studied, the oxytocin and vasopressin genes are located on the same chromosome locus, chromosome 20 in humans.6 Derived from a common ancestor, the oxytocin and vasopressin genes are small, encompass no more than 2.5 kilobases of genome, and have considerable homology. Each gene has three exons with two intervening sequences. Exon A exhibits 70% homology, while exon B has striking homology with few or no nucleotide differences, depending on the species. Exon C has the least homology between the genes for oxytocin and vasopressin.7 It appears that there are oxytocin enhancers localized within or in the vicinity of the vasopressin gene because oxytocin gene expression in transgenic mice will occur in magnocellular neurons only if the oxytocin gene is linked to the vasopressin gene in a minilocus.8 The oxytocin gene has three hormone-responsive elements: the estrogen-responsive element, the glucocorticoid-responsive element and the thyroid-responsive element. Thus, estrogen, glucocorticoids and thyroid hormone can regulate the oxytocin gene and its expression in several reproductive tissues in different species.
Vasopressin and oxytocin are synthesized as parts of longer independent proteins, preprohormones that include a second polypeptide entity called neurophysin. Oxytocin and vasopressin are synthesized initially as this preprohormone that is rapidly broken down to the active hormone and its neurophysin prior to its packaging with its respective neurophysin into neurosecretory granules. The neurophysin-hormone complex forms into distinct structures, the neurosecretory granules, which are distributed throughout the perikarya and along the neurons of the neurosecretory cells, whose axons pass through the median eminence into the neural lobe of the pituitary gland. The granules are transported down the axon at 1–3 mm/hour to reach the posterior pituitary gland.9, 10
The neurosecretory granules are released by exocytosis, and the membranes of the granules are recaptured by micropinocytosis.11 Thus, the neurohypophysial hormone and its neurophysin appear at the same time in the peripheral blood. Oxytocin is weakly bound to neurophysin and is, therefore, readily available for release in response to a stimulus. Data from Zimmerman et al. indicate that vasopressin and neurophysin not only are secreted in the neurohypophysial tract, but also are present in the hypophysial portal blood in rhesus monkeys, suggesting that vasopressin and oxytocin may affect anterior pituitary function.12
Oxytocin can appear in variable molecular forms with different biological properties for which transcriptional and posttranslational events play an essential role.13 Three types of oxytocin-related peptides, the COOH-terminally extended forms, Nα-acetylated oxytocin and oxytocin metabolites have been found. Their biological relevance is unclear.
REGULATORY MECHANISMS AND SECRETION
Vasopressin
Vasopressin is secreted from the neurohypophysial tract into the hypophysial portal blood and the cerebrospinal fluid. Three major stimuli control the release of vasopressin: (1) changes in osmolality of the blood; (2) alterations in blood volume; and (3) psychogenic stimuli, such as pain, fear, and apprehension. The osmoreceptors are located in the hypothalamus, while the volume receptors are found in the left atrium, aortic arch, and carotid sinus. Afferent impulses from the volume receptors travel via the vagus, glossopharyngeal, and aortic nerve to the diencephalon and decrease the secretion of vasopressin. Plasma osmolality changes are relatively more dominant than blood volume alterations in affecting vasopressin release in the human.14 Other compounds that may be involved in vasopressin release include: (1) angiotensin II, which produces a release of vasopressin, suggesting a feedback loop between the kidney and the hypothalamus; (2) cortisol, which may modify certain stimuli for the release of vasopressin, such as increasing the osmotic threshold for the release of vasopressin; and (3) atrial natriuretic factor (atriopeptin III), which inhibits the hyponatremia-stimulated release of vasopressin.15
Oxytocin
Two pathways for the peripheral regulation of oxytocin originate from areas in the nipple and the cervix. In the milk-ejection reflex, stimulation of the nipples triggers impulses which proceed along the peripheral nerves (thoracic 3, 4, and 5) and the spinal cord to the hypothalamus, and results in the release of oxytocin from the posterior pituitary gland. The released oxytocin then completes the reflex arc by stimulating contractions in the myoepithelial cells surrounding the alveoli and small ducts in the breast, resulting in milk ejection. The milk-ejection reflex can be suppressed by the activity of higher centers in the brain.
Cervical and vaginal stimulations result in the Ferguson reflex. This is mediated by impulses conducted from the peripheral nerves in the cervix and vagina to the spinal cord and up to the hypothalamoneurohypophysial system where oxytocin is released into the circulation and reaches the uterus to cause uterine contractions.
At a central level, factors controlling the release of neurohypophysial hormone are uncertain but may include: (1) adrenergic and cholinergic mechanisms in the hypothalamus; (2) neural impulses in the axons of the hypothalamoneurohypophysial tract; (3) local ionic changes in the region of the axon termina16 (in particular, release of vasopressin is calcium-dependent); (4) prostaglandins, which release oxytocin; (5) dopaminergic mechanisms (dopamine inhibits oxytocin release);17 (6) cholecystokinin18 and relaxin,19 both of which stimulate oxytocin secretion; and (7) thyrotropin-releasing hormone, which stimulates oxytocin and vasopressin release.20 The release of oxytocin has been shown to be episodic but is so rapid and frequent that it is preferable to describe it as “spurt” release. There are about three spurts of oxytocin release every 10 minutes;21 the spurts become more frequent and have greater amplitude during labor.2, 21, 22
Both oxytocin and vasopressin can increase the overall mitotic rate of acidophils in the adenohypophysis. It has been suggested that the neurohypophysial hormones might influence mitogenesis in the anterior pituitary either directly or by enhancing the secretion of the hypothalamic releasing factors.23 In humans, however, oxytocin infusion did not produce any change in circulating pituitary gonadotropins.24 Oxytocin modulates the central nervous system action or secretion of other neuropeptides and pituitary hormones. There is a high concentration of oxytocin in the hypophysial portal blood, and it is present in the neuronal elements of the external layer of the median eminence. Oxytocin appears to stimulate the release of prolactin25 and enhances thyrotropin-induced prolactin release during the normal menstrual cycle.26 Oxytocin acts reciprocally against vasopressin to inhibit its potentiating effect on corticotropin releasing factor-stimulated adrenocorticotropic hormone (ACTH)-cortisol release.27
ACTION AND METABOLISM OF NEUROHYPOPHYSIAL HORMONES
Both oxytocin and vasopressin have several specific receptors in the various target cells on which they act.28 Specific receptors for oxytocin have been demonstrated in cell membranes of the rat mammary gland, human uterine muscle, rat oviduct, corpus luteum, chorion, amnion, decidua, and the thymus and in multiple distinct brain regions (hypothalamus, hippocampus, olfactory system, limbic system, brain stem, certain parts of the striatum and cortex, the neurohypophysis, the ependyma of the lateral ventricle, and the choroid plexus near the lateral septum.)29 The oxytocin receptor present in human myometrium has a molecular weight of 43 kDa, consists of 388 amino acids, and has seven transmembrane domains typical of G-protein-coupled receptors.30 In human myometrium, endometrium, and ovary, the oxytocin receptors present are encoded by mRNAs with a 4.4 kilobase cDNA, while in the breast the mRNA is 3.6 kilobases.
Oxytocin stimulates smooth muscle contraction by a direct effect on the cell membrane through an increase in the number of normally sparse sodium gates. This action is mediated by cyclic adenosine monophosphate (cAMP) and guanosine monophosphate (cGMP). Oxytocin acts on its receptors in the myometrial cell to bring about two different effects. The first is opening the voltage-dependent calcium channels to increase intracellular free calcium concentration, which activates myosin light chain kinase and subsequent contractile steps. The second is oxytocin stimulation of inositol phosphate generation through phosphodiesterase cleavage of phosphatidyl inositol biphosphate.31 As a neuropeptide, oxytocin exerts a neuromodulatory effect on the central nervous system through its receptors present in the brain. This neuromodulatory effect of oxytocin is involved in and affects drug addiction, ethanol tolerance, learning and memory, sexual and maternal behavior,32 and inhibition of food intake,33 and plays a role in various forms of social dysfunction—autism, social anxiety disorder, borderline personality, and schizophrenia.34
Vasopressin acts differentially in various target tissues through three distinct G-protein receptors identified by second messenger and variable affinity for prepared analogues.28 V1 (previously V1A) receptor is identified in liver, vascular smooth muscle, and most peripheral tissues, while V3 (previously V1B)35 is found in adenohypophysis, and V2 is found in kidney. V1 and V3 receptors are coupled to phospholipase C, while the vasopressin V2 receptor utilizes adenylate cyclase which catalyzes the conversion of adenosine triphosphate (ATP) to cAMP. It is not clear how an increase in cAMP leads to the antidiuretic response, but it is suggested that by activating a protein kinase that catalyzes phosphorylation of membrane proteins, the permeability of the membrane is altered to increase water diffusion. Vasopressin also does the following: (1) stimulates sodium diffusion; (2) stimulates smooth muscle contraction in the vascular bed, leading to the observed pressor action seen after physiologic and pharmacologic doses of vasopressin; (3) stimulates the anterior pituitary to cause release of ACTH after large doses of vasopressin; (4) stimulates directly the release of corticotropin-releasing factor; and (5) enhances memory. The high levels of vasopressin found in the hypophysial portal blood could play a part in the release of ACTH.
Both oxytocin and vasopressin circulate as free peptides since the affinity constant of neurophysins is too low to permit significant binding. Data on the apparent volume of distribution of oxytocin indicate that it is apparently distributed into both the intravascular and extravascular compartments. Both neurohypophysial hormones are cleared by the liver and the kidneys. The half-life of vasopressin is 3–6 minutes. The half-life of oxytocin is 5–17 minutes, with a mean of 10 minutes. The half-life of oxytocin is not significantly reduced during pregnancy, but it is reduced if the dose of oxytocin infused is increased.24, 36, 37 The metabolic clearance rate of oxytocin is 21.5 ml/kg/min and is not affected by pregnancy.24 During pregnancy, oxytocin is also metabolized in the circulation and in the placenta by the placental enzyme oxytocinase, which is an aminopeptidase, the concentration of which increases with advancing gestation. Oxytocinase cleaves the link between the N-terminal hemicystine residue (position 1) of oxytocin and the adjacent tyrosine residue (position 2), destroying the ring structure and biologic activity of the molecule. The peptide chain is further cleaved in succession up to the proline residue at position 7. Degradation of oxytocin by oxytocinase may be a protective mechanism against the uterine-stimulating effects of excessive circulating oxytocin, but the failure of pregnancy plasma to reduce the half-life of oxytocin both in vivo and in vitro argues against a major role of oxytocinase in the metabolism of oxytocin during pregnancy.
METHODS OF MEASUREMENT
Oxytocin and vasopressin can be measured either by biologic assays or by radioimmunoassays. The bioassays of vasopressin have been based on its antidiuretic action on a hydrated ethanol-anesthetized rat or on its effects on smooth muscle. The bioassays of oxytocin are based on the milk-ejection reflex as measured by the mammary intraductal pressure or the response of uterine muscle strips in vitro. These assays are relatively insensitive, require large volumes of blood, and are susceptible to poor specificity. Many biologically active substances, such as bradykinin, angiotensin, acetylcholine, serotonin, and histamine can interfere with the bioassay.
In view of these problems, radioimmunoassays of oxytocin and vasopressin have been developed by several investigators.38, 39, 40, 41 Although radioimmunoassay of vasopressin and oxytocin is available in some laboratories, its use is still confined to research investigations because of the limited availability of antiserum and the technical difficulties of the assay. Because of the low circulating concentrations of neurohypophysial hormones and the interference with antigen-antibody binding by nonspecific factors in the plasma, many of the radioimmunoassay methods that have given apparently reliable results have employed methods to extract the neurohypophysial hormones from plasma prior to measurement. The advantages of radioimmunoassay over bioassay of oxytocin and vasopressin include high specificity, precision, and reliability, as well as increased sensitivity. The antisera used in many reported studies are able to accurately distinguish vasopressin from oxytocin.36, 38, 39 Some dissociation between the values of neurohypophysial hormones measured by radioimmunoassay and bioassays has been found, but in most instances the results obtained between the two methods of assay seem to show satisfactory correlation.30, 42
Neurophysins I and II have been measured by radioimmunoassay only. Oxytocin, vasopressin, and their neurophysins can be characterized and also measured by high-pressure liquid chromatography. While separation and characterization are readily attained by high-pressure liquid chromatography, radioimmunoassay is more sensitive for quantification.
TESTS OF POSTERIOR PITUITARY FUNCTION
All tests of posterior pituitary function are based on the release or suppression of vasopressin. There is no test that will give precise information on oxytocin reserve in the posterior pituitary gland. The posterior pituitary function tests have been used to distinguish among cranial diabetes insipidus, nephrogenic diabetes insipidus, and psychogenic polydipsia. The tests available include water deprivation tests, the hypertonic saline infusion test, and the nicotine stimulation test.
Water deprivation tests
Many different types of the water deprivation test have been used. A properly conducted 8-hour water deprivation test is adequate. In normal subjects, the urine osmolality is greater than 800 mOsm/kg and plasma osmolality is less than 294 mOsm/kg at the end of the 8 hours of fluid deprivation. In diabetes insipidus, the urine osmolality is less than that of the plasma, which is usually greater than 300 mOsm/kg at the end of the 8 hours. Patients with nephrogenic diabetes insipidus can easily be distinguished from those with cranial diabetes insipidus since they do not respond to exogenous vasopressin (desmopressin, 20 μg), which is given at the end of the water deprivation with urine collection continued for another 4 hours. Patients with psychogenic polydipsia usually have normal or low plasma osmolality before the test and have an impaired ability to concentrate their urine. If it can be measured, urinary immunoreactive vasopressin in patients with cranial diabetes insipidus shows a minimal rise with fluid deprivation as opposed to a normal rise in patients with nephrogenic diabetes insipidus.
The other posterior pituitary function tests (hypertonic saline infusion and nicotine stimulation tests) have been introduced to overcome the potential pitfalls of the water deprivation test, since in localized hypothalamic disease the osmolar release of vasopressin may be affected while the volume-mediated control mechanisms are unaffected. Some workers believe, however, that the water deprivation test can identify hypothalamic disease affecting vasopressin release.
Hypertonic saline infusion test
When hypertonic saline is infused in volunteers or patients the renal–vasopressin axis is stressed. Those with normal osmoregulation will mount a response of increased circulating vasopressin and maintain normal plasma osmolality.43 Patients with central (cranial) diabetes insipidus (DI) will develop increased plasma osmolality and fail to concentrate the urine to retain free water in order to normalize the osmolality. Plasma vasopressin levels measured during the infusion also fail to rise. This test will differentiate patients with central DI from those who have a more normal vasopressin secretion response to increased plasma osmolality—identifying primary polydipsia or nephrogenic DI.
Nicotine stimulation test
The nicotine stimulation test is less well-established than the water deprivation test and requires different dosages of nicotine for smokers (up to 3 mg) and nonsmokers (1 mg). The mechanism of nicotine-mediated vasopressin release may be via both the cholinergic innervation of the neurohypophysis and the result of hypotension, with accompanying reduction in glomerular filtration rate and urine flow rate.
OXYTOCIN
Direct evidence has shown that oxytocin is involved or mediates several events in reproductive physiology.
Female
DURING COITUS
Oxytocin is released during coitus, but the stimuli responsible for the release are not clearly known. It may involve olfactory, visual, and auditory pathways, or it may be an expression of the Ferguson reflex as a result of vaginal and cervical stimulation during coitus. Circulating oxytocin levels increase significantly with sexual arousal and at ejaculation in males during masturbation,44 as well as during coitus.36 In women, maximum levels of oxytocin are found at the peak of orgasm.45, 46 Contractions induced by oxytocin release at orgasm may facilitate sperm transport through the uterus, enhancing reproductive potential through increased sperm–oocyte interaction, although this concept has been challenged.47
DURING PREGNANCY
Few studies have measured oxytocin in the blood or amniotic fluid throughout pregnancy. Using a specific and sensitive radioimmunoassay, it was found that oxytocin is detectable in 85% of the maternal plasma taken from pregnant women at 6–42 weeks' gestation, while amniotic fluid has detectable oxytocin levels in 90% of the samples studied.21 Maternal plasma oxytocin increases significantly with gestational age. The mean maternal plasma oxytocin level increases from 10.4 pg/ml at 8–9 weeks to 26.4 pg/ml at 37 weeks and is followed by a significant further increase to 74.2 pg/ml at 38 weeks.21 The increase in maternal plasma oxytocin throughout pregnancy may be due to the rise in circulating estrogen levels, since the administration of estrogens in adult males stimulates oxytocin release. The rising levels of circulating progesterone throughout pregnancy until term could be responsible for blocking the effect of oxytocin on an increasingly estrogen-primed uterus and, thus, prevent premature onset of uterine contractions. The maternal pituitary is the primary source of the circulating maternal oxytocin during pregnancy, although oxytocin gene activity is demonstrated by detection of oxytocin mRNA in the decidua,48, 49 indicating that it may be an additional source of oxytocin during pregnancy. Moreover, the close proximity of the decidua to the uterine myometrium makes it a likely candidate for paracrine activity at term, possibly initiating or augmenting oxytocin stimulation of myometrial contractions.
Amniotic fluid oxytocin concentration increases from a mean of 7.8 pg/ml at 14–15 weeks' gestation to 43.9 pg/ml at 40 weeks and 30.8 pg/ml at 41–42 weeks. Fetal urine has been shown to contain oxytocin with a mean level of 54 pg/ml50 and may contribute substantially to the amniotic fluid concentration. An amniotic fluid to fetal urine oxytocin concentration ratio of 2:1 in spontaneous labor indicates that there are other sources of amniotic fluid oxytocin.50 Direct efflux of oxytocin from cord vessels is a possible source. Meconium, which is rich in oxytocin,51 contributes to amniotic fluid oxytocin in special circumstances in which enhanced uterine activity may be necessary to hasten delivery of a fetus that is in distress. Since mRNA for oxytocin has been found in human amnion, chorion, and decidua, these are also sources of amniotic oxytocin.48, 49 With radioimmunoassay, rather high concentrations of circulating oxytocin have been found by some investigators52 who do not use preliminary extraction of the plasma in their methods compared with those who do.36, 38
DURING LABOR AND PARTURITION
Data available from the literature on the maternal plasma oxytocin levels during human labor are summarized in Table 1. Coch and colleagues53 found significantly higher concentrations of oxytocin in the jugular venous blood compared with peripheral blood and concluded that the maternal pituitary gland releases significant amounts of oxytocin during labor. Chard and colleagues22, 36 found detectable oxytocin levels in 19–60% of the maternal peripheral blood samples during labor. It is believed that the absence of detectable oxytocin in some labor samples is due to spurt release of oxytocin. They found that the percentage of blood samples with detectable levels of oxytocin rises in the late first and second stages of labor. Dawood, Raghavan, and Pociask found that maternal plasma oxytocin increases significantly from the first stage to the second stage of labor, followed by an equally significant decline during the third stage of labor (Figure 3).38, 54
Table 1. Cumulative data on oxytocin concentration in maternal blood during labor
| Author | Stage of labor | Source of blood assayed | Method of assay | Oxytocin levels |
| Hawker & Robertson (1957) | First | Arm vein | Bioassay with extraction | 200–2400 pg/ml blood |
| Caldeyro-Barcia (1961) | First | Arm vein | Bioassay without extraction | 240 pg/ml plasma |
| Hawker et al (1961) | First | Arm vein | Bioassay with extraction | 0–11,900 pg/ml blood |
| Hawker et al (1961) | Second stage (expulsive phase) | Arm vein | Bioassay without extraction | 280 pg/ml blood |
| Juret et al (1961) | First stage | Arm vein | Bioassay without extraction | 60,000–100,000 pg/ml plasma |
| Coch et al (1965)
| First, second, third | Jugular vein | Bioassay with extraction | 600–1800 pg/ml plasma |
| First, second, third | Arm vein | Bioassay with extraction | 50–500 pg/ml plasma | |
| Fitzpatrick & Walmsley (1965) | Second | Arm vein | Bioassay with extraction | 160–400 pg/ml plasma |
| Vorherr (1977) | First, second | Arm vein | Bioassay | Undetectable |
| Saameli (1963)
|
|
| Indirect assessment using oxytocin intravenous infusion
| 6 pg/ml plasma
|
| Chard et al (1970) |
| Arm vein | RIA with extraction | Detectable levels in 19% |
| Gibbens & Chard (1976) | First | Arm vein | RIA with extraction | Detectable levels in up to 60%* |
| Bashore (1972)
| First, second, third
| Arm vein
| RIA with extraction
| <40 pg/ml plasma |
| 180 pg/ml plasma (2 cases) | ||||
| Kumaresan (1974) | First, second | Arm vein | RIA without extraction | 70–870 pg/ml plasma |
| Glick et at (1969)
| First, second
| Arm vein
| RIA with extraction
| 20–150 pg/ml plasma |
| 200–400 pg/ml plasma | ||||
| Dawood et al (1978)
| First, second, third (expulsive phase) | Arm vein | RIA with extraction
| 40.3 ± 9.8 pg/ml plasma† |
| 123.9 ± 23.6 pg/ml plasma | ||||
| 64.5 ± 13.1 pg/ml plasma |
*Oxytocin levels given are 0–25 pg/ml.
† Mean ± standard error of the means.
RIA, radioimmunoassay.
(Modified from Dawood MY: Contemp Obstet Gynecol 13:181, 1979.)
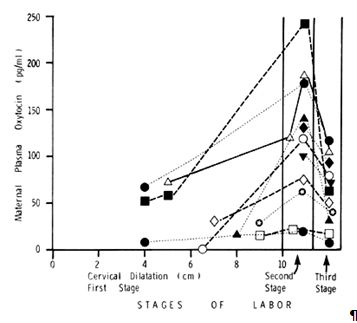
Fig. 3. Serial maternal plasma oxytocin during the first, second, and third stages of labor in 11 women with spontaneous normal labor and vaginal delivery. In every patient, there was an increase in plasma oxytocin from the first to the second stage and a fall from the second to the third stage. The plasma samples taken during the second stage of labor were timed to coincide with the crowning of the fetal head. (Modified from Dawood MY, Raghavan KS, Pociask C, Fuchs F: Oxytocin in human pregnancy and parturition. Obstet Gynecol 51:138, 1978)
During the first stage of labor, the mean maternal plasma oxytocin level is 40.3 ± 9.8 pg/ml. It increases to 123.9 ± 23.6 pg/ml during the second stage and declines to 64.5 ± 13.1 pg/ml during the third stage. This occurs both in the individual patient and in all the patients studied as a group. Hence, the release of oxytocin into the maternal circulation during the second stage is accentuated during the passage of the presenting part through the cervix and the vaginal outlet via the mechanism of the Ferguson reflex. Because higher levels of oxytocin are found in the jugular venous blood than in the peripheral blood,53 the maternal posterior pituitary gland must be releasing oxytocin during labor to increase the peripheral maternal oxytocin levels.
FETAL OXYTOCIN
As determined in umbilical cord blood samples, irrespective of the mode of delivery, fetal plasma oxytocin concentrations are higher when there is spontaneous onset of labor compared with oxytocin levels in samples taken from infants of women who are not in labor when undergoing cesarean section.36, 50 Dawood et al. found that the plasma oxytocin levels are always higher in the umbilical artery than in the umbilical vein whether women are in labor or not, provided they are not given oxytocin. When women are in established labor, the umbilical arterial plasma oxytocin levels are significantly higher than the umbilical venous plasma oxytocin levels (Figure 4). Because the oxytocin levels are always higher in the umbilical artery than in the umbilical vein, regardless of the stage of labor and fetal presenting part, the oxytocin must originate in the fetus with the flow from the fetal side toward the maternal compartment. When patients are given intravenous or buccal oxytocin to stimulate uterine contractions, levels in the umbilical vein are higher than or similar to the levels in the umbilical artery. This finding indicates that it is possible to induce a reverse gradient of oxytocin toward the fetal compartment. This suggests that in pregnant patients requiring oxytocin stimulation of the uterus, there is an underlying deficiency in the release of oxytocin by the fetus and possibly by the decidua, chorion, or amnion, which accounts for the need for oxytocin administration.
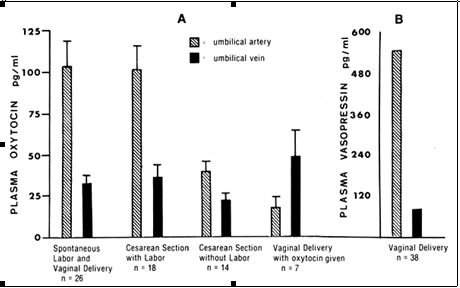 Fig. 4. A. Oxytocin levels in the umbilical arterial and venous blood during elective cesarean section, during cesarean section after the onset of labor, and on administration of oxytocin to the mother. Note that umbilical arterial plasma always had higher oxytocin levels that umbilical venous plasma except when the mother was given oxytocin. The umbilical arteriovenous difference in oxytocin levels is significantly higher when there is spontaneous labor than when there is no labor. (Data from Dawood MY, Wang CG, Gupta R, Fuchs F; Fetal contribution of oxytocin in human labor. Obstet Gynecol 52:205. 1978. Vasopressin data not originally published)
Fig. 4. A. Oxytocin levels in the umbilical arterial and venous blood during elective cesarean section, during cesarean section after the onset of labor, and on administration of oxytocin to the mother. Note that umbilical arterial plasma always had higher oxytocin levels that umbilical venous plasma except when the mother was given oxytocin. The umbilical arteriovenous difference in oxytocin levels is significantly higher when there is spontaneous labor than when there is no labor. (Data from Dawood MY, Wang CG, Gupta R, Fuchs F; Fetal contribution of oxytocin in human labor. Obstet Gynecol 52:205. 1978. Vasopressin data not originally published)
It is still unclear whether all the oxytocin flowing from the fetal side into the maternal compartment crosses the placenta without being inactivated by the high concentrations of placental oxytocinase. It is clear, however, that an oxytocin gradient can be established from the fetus to the mother or from the mother to the fetus and that oxytocin readily crosses the placenta in the ewe55 and the baboon.56 A major role of oxytocinase in the metabolism of oxytocin has been seriously questioned (see section on Metabolism). Since the uterus is the principal tissue with specific oxytocin receptors and is immediately receiving the fetal hormones after its transplacental passage, much, if not all, of the fetal oxytocin in the umbilical artery must be readily available to stimulate the uterus. Thus, the fetus appears to act as an “oxytocin-injection” system to the mother. Additional sources of oxytocin such as the decidua, chorion, and amnion can also act in a paracrine manner on the uterus.48
The umbilical plasma arteriovenous difference in oxytocin concentration is significantly higher when the woman is in established labor than when she is not.50 Using the arteriovenous difference in umbilical plasma oxytocin levels, the average umbilical blood flow, and umbilical blood hematocrit levels, the estimated mean secretion rates of oxytocin from the fetal to the maternal compartments have been shown to be 2.75 mU/min (5.5 ng) when there is spontaneous labor and vaginal delivery; 3 mU/min (6 ng) in patients having cesarean section after the onset of labor; and only 0.5 mU/min (1 ng) if cesarean section is performed without onset of labor (Table 2).50 It is noteworthy that the secretion rate of 0.5 mU/min in women who are not in labor is consistent with the clinical observation that only ineffectual uterine contractions, if any, will occur with the administration of such a small dose of oxytocin, but that infusion rates of oxytocin similar to the secretion rate of 2.75–3 mU/min found in spontaneous labor will cause effective uterine contractions at term when there has been an increase in oxytocin receptors. Since the secretion rate of oxytocin from the fetus to the mother during the first stage of labor, as shown in women undergoing cesarean section with labor, is similar to that found during the second stage in spontaneous labor and vaginal delivery, the sudden increase in maternal plasma oxytocin observed at the second stage must be due to additional oxytocin released from the maternal neurohypophysis. Hence, the data available indicate that during the first stage of spontaneous labor, fetal oxytocin plays a significant and major role, while the maternal pituitary releases additional oxytocin to increase markedly the maternal circulating oxytocin concentration during the second stage (expulsive phase). This increase is necessary to augment maternal oxytocin levels already present in the first stage of labor and induce the strong uterine contractions required to expel the fetus during the final stage of its exit from the birth canal.
Table 2. Estimated secretion rate of oxytocin from the fetal to maternal compartment based on umbilical plasma arteriovenous differences in oxytocin concentrations
|
|
(Data from Dawood MY, Wang CF, Gupta R, Fuchs F: Fetal contribution to oxytocin in human labor. Obstet Gynecol 52:205, 1978.)
Fetal urine has been shown to contain oxytocin.50 First-voided fetal urine obtained after delivery has a mean oxytocin concentration of 53.8 ± 11.8 pg/ml, a higher level than that reported by Boyd and Chard in maternal urine during labor.57 This provides further evidence for fetal participation in oxytocin release during spontaneous labor.
Little is known about the ontogeny of human neurohypophysial hormones. The concentration of oxytocin in the human posterior pituitary gland of 150–156-day-old fetuses was determined by bioassay to be 92–105 ng/gland,.58] Using radioimmunoassay, Khan-Dawood and Dawood found that at 14–17 weeks' gestation, fetal pituitary gland had 10.2 ± 5.9 ng oxytocin/gland (mean ± SEM), increasing to 31.6–38.4 ng at 20–26 weeks; 22.1–57 ng at 32 weeks; and dramatically to 544.3 ± 33.8 ng oxytocin/gland in 1–5-day-old newborns.59 Thus, it is evident that the fetal pituitary has started to produce oxytocin by the 14th week of pregnancy, if not earlier.
Vasotocin is present in the human fetal posterior pituitary gland. Extracts of human fetal neurohypophysial tissue at 130–155 days' gestation had oxytocin, vasopressin, and vasotocin. Based on tissue cultures of fetal neurohypophysial cells, it has been shown that vasotocin is produced and secreted by the ependymal cells.60 Thus, while oxytocin and vasopressin are synthesized in the hypothalamic neurosecretory system, vasotocin, which has been suggested as the ancestral molecule, is synthesized in the ependymosecretory system, which lacks neurons. The fetal posterior pituitary gland not only is the site of storage of neurohypophysial hormones but it also synthesizes vasotocin.
The precise stimulus that causes the release of oxytocin in the human fetus has not been identified, and it is not known whether the release is a primary phenomenon or secondary to the process of labor. It may be hypothesized that stress on the fetus may cause a release of neurohypophysial hormones, but the nature of the stress is difficult to define. It is possible that with increasing growth of the fetus, the placenta is unable to cope with the nutritional demands of the fetus and hence the fetus is stressed. On the other hand, activation of the fetal neurohypophysis during labor and delivery may be the result of acute stimulation caused by uterine contractions to a mature fetal hypothalamoneurohypophysial system.
IN PREMATURE LABOR
Ethanol can suppress uterine activity in premature human labor. It is unclear, however, whether the alcohol affects the myometrium, the posterior pituitary gland, or both. Evidence points to a posterior pituitary gland mechanism because the percentage of detectable oxytocin decreases significantly when alcohol is given during the advanced phase of the first stage of labor.17 Ethanol can also inhibit prostaglandin-induced uterine activity. It is conceivable that the abnormality due to endocrine factors in premature labor may be a deviation from the delicate balance between oxytocin, estrogens, and progesterone, each capable of influencing uterine activity directly or indirectly.
Other disorders associated with labor
There is no evidence of abnormal labor in patients with diabetes insipidus, although it might be expected that the onset of labor would be delayed in such patients if the maternal release of oxytocin is crucial. A surge in plasma oxytocin during labor and the puerperium, similar to the surge found in normal pregnancy, has been reported in a patient with idiopathic diabetes insipidus.61 Since this disease usually limits the function of the maternal posterior pituitary gland, the increase in oxytocin concentration in the maternal circulation is probably of fetal, decidual, chorionic, and/or amniotic origin and may account for the ability of these patients to go into spontaneous labor. In patients with idiopathic diabetes insipidus, all of the posterior pituitary gland may not have been destroyed and the oxytocin reserve in most of the patients is unknown.
In a study on a limited number of patients, sequential maternal plasma oxytocin concentrations appear to correlate well with the subsequent outcome of labor.21 Patients who have plasma oxytocin concentrations of more than 10 pg/ml throughout the second half of their pregnancies (good oxytocin secretors) develop good rhythmic uterine contractions when they go into labor and deliver vaginally without any difficulty. By contrast, those who have plasma oxytocin concentrations of less than 10 pg/ml (poor oxytocin secretors) develop uterine dysfunction during labor, which necessitates cesarean section.
DURING LACTATION
Increased oxytocin concentrations are found in the maternal blood during suckling (Table 3). When the human infant suckles, rhythmic changes in intramammary pressure are evoked in the contralateral breast. These changes are caused by rhythmic contraction of the mammary myoepithelium in response to oxytocin. The release of oxytocin in response to suckling is mediated through impulses generated at the nipple and transmitted via the third, fourth, and fifth thoracic nerves to the spinal cord up to the hypothalamus. Hypothalamic responses generate release of oxytocin from the posterior pituitary, and the subsequent rise in circulating oxytocin levels results in increased binding of its receptors on the breast myoepithelium. This completes the milk-ejection reflex.
The critical role of oxytocin in lactation has been demonstrated in mice genetically engineered for oxytocin deficiency.62 Although oxytocin has recognized roles in sexual and reproductive behaviour, and a role in stimulating or augmenting contractions of the uterus at parturition, oxytocin deficient mice are viable and reproduce normally. Postpartum, oxytocin deficient females demonstrate normal maternal behavior, but all offspring die due to malnutrition when the mother fails to lactate or nurse.
In addition to milk ejection, a concurrent, parallel reflex arc is also responsible for the uterine contractions, often referred to as the “after pains,” observed at the time of breastfeeding. Using radioimmunoassay, Dawood et al. found that maternal circulating oxytocin levels are low, usually less than 30 pg/ml prior to suckling. The oxytocin level increases 5–10 minutes after initiating suckling, but the maximum levels reached are less than 100 pg/ml during a 30-minute suckling episode (15 minutes on each breast) in the first few days after delivery.63 In many instances, two peaks in the plasma oxytocin concentrations are observed, perhaps a reflection of the suckling stimuli induced in the contralateral breast when the infant is changed to the opposite side. The release of oxytocin during lactation may not be exclusively due to the suckling stimulus, but may include olfactory, auditory, and other stimuli. Thus, a mother who is breastfeeding may start to lactate upon hearing her infant’s cry. More information is needed on the relationship of the neurohypophysis to the adenohypophysis, the influence of oxytocin on prolactin release, and their respective participation in the mechanism of lactation.
Table 3. Maternal oxytocin levels during lactation
| Authors | Source of blood | Relationship to suckling
| Method of oxytocin determination
| Oxytocin level |
| Hawker & Robertson (1957)
| Arm vein
| Before suckling, during suckling
| Bioassay with extraction
| 1.4 ng/ml blood |
| <0.86 ng/ml blood | ||||
| Hawker (1958)
| Arm vein
| Before, during, and 2 hr after suckling
| Bioassay with extraction | 40 pg/ml blood |
| 100 pg/ml blood | ||||
| 8 pg/ml blood | ||||
| Hawker et al (1961) | Arm vein | Before suckling | Bioassay with extraction | 0–8.8 ng/ml blood |
| Coch et al (1965) | Internal jugular vein | During suckling | Bioassay with extraction | 400–600 pg/ml plasma |
| Coch et al (1968) | Internal jugular vein | During suckling | Bioassay with extraction and chromatography | 24–50 pg/ml plasma |
| Fox & Knaggs (1969) | Arm vein | Before suckling, during suckling | Bioassay with extraction
| 38 pg/ml plasma |
| 22–224 pg/ml plasma |
(Modified from Dawood MY: Contemp Obstet Gynecol 13:181, 1979.)
NIPPLE STIMULATION TEST
Clinically, the reflex arc, nipple–neurohypophysis–uterus, has been utilized to develop the Nipple Stimulation Test, a modification of the Oxytocin Challenge Test (OCT) or Contraction Stress Test. Initially described in an oral presentation at the 1980 ACOG District VIII meeting, MacMillan subsequently published his experience and numerous additional studies followed, including direct comparisons to the OCT.64, 65, 66 For the nipple stimulation test, women near term requiring fetal evaluation are monitored using tocodynamometry. The nipples are alternately rubbed with a warm wash cloth until uterine contractions develop. When three or more contractions in a 10 minute period develop, the fetal heart rate response is interpreted to determine the state of fetal well-being. The test offers the advantage of stress testing without a requirement for intravenous access. The brief popularity of this testing has been supplanted by Non-Stress Testing combined with ultrasound (biophysical profile).
Nipple stimulation has also been studied for the ability to shorten gestation67 or augment labor68, 69, with mixed results.
Neurohypophysial hormones in peripheral tissues
Oxytocin and vasopressin have been identified in corpora lutea of women70, 71, 72 and in human testes.73 Significantly higher concentrations of oxytocin are found in the corpus luteum at the midluteal phase compared to early and late luteal levels (Figure 5).72, 74 Ovarian vein blood draining from the region of corpora lutea has a higher concentration of oxytocin than when there is no corpus luteum,72, 74, 75, 76 and the mRNA for oxytocin is present in corpora lutea.73 Although the role of oxytocin in the ovary and testis is not well understood, oxytocin probably exerts an antigonadal effect and serves as an intragonadal modulator of luteal function.72, 75 Oxytocin inhibits in vitro luteal cell progesterone production.75 It shortens the luteal phase and reduces progesterone levels when given directly into the corpus luteum of monkeys.77 Oxytocin is also present in human adrenal glands.78 In the adrenal gland, oxytocin appears to regulate steroidogenesis. Oxytocin present in peripheral nonneural tissues likely exerts a paracrine or autocrine effect.
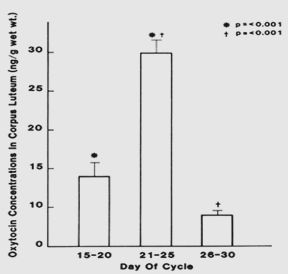 Fig. 5. Oxytocin concentrations in human corpus luteum during the early (days 15–20) and late (days 26–30) luteal phase show a significant increase at midluteal phase followed by a significant decrease in the late luteal phase. (Dawood MY, Khan-Dawood FS: Human ovarian oxytocin: Its source and relationship to steroid hormones. Am J Obstet Gynecol 154:756, 1986
Fig. 5. Oxytocin concentrations in human corpus luteum during the early (days 15–20) and late (days 26–30) luteal phase show a significant increase at midluteal phase followed by a significant decrease in the late luteal phase. (Dawood MY, Khan-Dawood FS: Human ovarian oxytocin: Its source and relationship to steroid hormones. Am J Obstet Gynecol 154:756, 1986
Male
In the male, the function of oxytocin, if any, is unclear. Oxytocin is released and increases significantly with sexual arousal and also at ejaculation both with masturbation44 and with coitus.45 Several studies have shown that exogenous oxytocin may affect sperm transport, possibly by an action on the smooth muscle in the seminiferous tubules. In male rabbits, administration of methallibure, an inhibitor of oxytocin release, reduces sperm count markedly.79 This effect is abrogated when oxytocin is given simultaneously with methallibure, suggesting that oxytocin released during ejaculation may play a part in sperm transport.
Oxytocin is present in the testes of several species. In rather high pharmacologic doses, oxytocin exerts an antigonadal effect on the testes by inhibiting testosterone output.80
VASOPRESSIN
Adult
Unlike oxytocin, which is released in spurts, vasopressin appears to be secreted continuously in normal hydrated humans. The basal levels, as determined by both radioimmunoassay and bioassay, range from 1 to 5 pg/ml. The plasma vasopressin levels after dehydration range from 1.3 to 16.4 pg/ml. Some workers found no effect of posture on plasma vasopressin, while others found a significant increase in plasma vasopressin in the erect posture.
In patients with cranial diabetes insipidus, the mean plasma vasopressin level is about 3.5 pg/ml (range, 1.4–5 pg/ml) and is significantly lower than in normal subjects. These patients fail to increase their plasma vasopressin following fluid restriction.
In patients who have the syndrome of inappropriate antidiuretic hormone (SIADH), the reported level of plasma vasopressin is 3.2–22 pg/ml. A striking feature of this disease is that in patients with SIADH due to an oat cell bronchial carcinoma, plasma vasopressin levels are appreciably higher (13–283 pg/ml) than in SIADH not due to an oat cell bronchial carcinoma.
Fetus
While there is little information available on the maternal vasopressin levels in humans, the umbilical arterial and venous plasma levels of vasopressin from infants experiencing labor have been shown to be quite high when compared to values from non-labored infants, and 5–15 times higher than the oxytocin concentration (Figure 3). Like oxytocin, vasopressin levels are higher in the umbilical artery than in the vein at delivery. The release of such large amounts of vasopressin may be a reflection of stress or hypoxia on the fetal neurohypophysis at the time of parturition but remains to be fully elucidated. The significance and purpose of the high concentrations of vasopressin at the time of delivery are not understood. Several suggestions may be advanced. Vasopressin can stimulate uterine contractions, although it is much less potent than oxytocin in this regard. When the 10–15 fold higher concentration of vasopressin than oxytocin in the umbilical artery is considered, however, the amount of vasopressin in the umbilical artery may be almost equipotent to the amount of oxytocin in terms of eliciting uterine contractions.
Vasopressin can stimulate the release of adrenocorticotropin hormone (ACTH) from the adenohypophysis. This may be part of the mechanism responsible for the physiologic stimulation and release of ACTH in the fetus at the onset of labor leading to a rise in fetal cortisol that occurs at the same time.
In the newborn, release of vasopressin in response to severe hypoxia or during the respiratory distress syndrome can contribute to the abnormal renal function that may accompany this condition.
LEVELS OF NEUROPHYSIN
It has been suggested that since measurement of vasopressin and oxytocin is quite difficult, more easily measured neurophysin levels might reliably reflect oxytocin and vasopressin release. Evidence fails to bear this out; first, the binding affinity of neurophysin to the neurohypophysial hormones at the pH of blood in the human is so low that little oxytocin is bound to neurophysin. Second, the metabolic clearance rate of neurophysin, which has a molecular weight 10 times greater than that of the neurohypophysial hormones, may be significantly different from that of oxytocin and vasopressin, leading to marked differences in calculated circulating concentrations of each.
In adult humans, the circulating levels of neurophysin decrease with age.81 The reason for this is unclear. During the menstrual cycle, neurophysin I and neurophysin II levels increase during the follicular phase of the cycle but the level of neurophysin I is constantly higher than that of neurophysin II.81 The increase in the level of neurophysin I has been attributed to stimulation of its release by estrogens, which increases with follicular development and maturation. Hence, it appears that estrogen may modulate neurohypophysial function in the human. In adult males, administration of estrogen can cause a release of neurophysin I.
During pregnancy, there is a gradual increase in immunoreactive neurophysins in the circulation but there is a wide dispersion of individual values.81 In contrast to the follicular phase of the menstrual cycle, neurophysin II is always higher than neurophysin I throughout pregnancy, although circulating estrogen levels are rising substantially throughout pregnancy. Circulating neurophysin levels do not show any significant change during suckling, parturition, or sexual intercourse.82 This finding is again seemingly contrary to expectation if the neurohypophysial hormones are involved in these processes, but may be understood if the binding and metabolic properties of neurophysins are considered.
While the adult pineal gland does not have neurophysin, the fetal pineal gland has been found to have a neurophysin-like substance. It has been suggested that this may represent the physiologic gonadotropin-inhibiting activity of the pineal gland. Neurophysin has also been detected in the anterior pituitary gland, although in concentrations 100 times less than in the posterior pituitary gland. These observations together with the finding of high concentrations of neurophysins and vasopressin in the hypophysial portal circulation of rhesus monkeys12 suggest that there is an interrelationship between the anterior and posterior pituitary glands, the exact nature of which remains to be elucidated.
The levels of neurophysin in the infant are higher than in the adult but decrease with age. Neurophysin levels are higher on the first day of life than on the sixth day, and neurophysin II levels are always higher than those of neurophysin I in the neonate.
Neurophysins are also found in non-pituitary tissues. Neurophysin I and II are found in the kidney cortex and the mammary gland, but only neurophysin I is found in the pineal gland. The significance of this observation is unclear. Neurophysins have also been immunocytochemically localized in the corpora lutea of women and baboons.83, 84
PHARMACEUTICAL ACTIVITIES AND THE NEUROHYPOPHYSIAL HORMONES
Drugs that stimulate the release of vasopressin include acetylcholine, nicotine, α-agonists given directly into the cerebral ventricles, β-agonists given systemically, vincristine, bradykinin, and clofibrate.
Ethanol, α-agonists, and atropine given via the systemic circulation have been used successfully to induce diuresis in patients with syndrome of inappropriate antidiuretic hormone. The mechanism appears to be inhibition of release of vasopressin, but they are effective only when there is a benign cause for the syndrome.
A powerful mediator of vasopressin activities is 1-desamino-8-D-arginine vasopressin, desmopressin or DDAVP. Due to the deamination of amino acid 1 and the dextro form of arginine substituted at position 8, this synthetic peptide is degraded more slowly than the native or recombinant forms of vasopressin. Acting via the V2 receptors in the kidney, DDAVP increases water reabsorption. In the management of central DI, DDAVP can be administered to replace absent vasopressin. It may be used in the evaluation of patients with DI to differentiate the central (cranial) from the nephrogenic form of the disease. This agent is utilized to reduce free water clearance during sleep in the treatment of nocturnal enuresis or nocturia associated with sleep apnea. It also mediates release of von Willebrand factor by endothelial cells and promotes clotting by increasing factor VIII survival due to complexing of the von Willebrand factor. This coagulation enhancement is beneficial in von Willebrand disease, mild hemophilia A (factor VIII deficiency), and thrombocytopenia.
DDAVP is available in intravenous, intramuscular, subcutaneous, intranasal, sublingual, and oral forms. The intranasal forms have been banned by the Food and Drug Administration for treatment of sleep apnea after two patients died and 59 others suffered seizures. DDAVP tablets are still considered safe for treatment of sleep apnea in otherwise healthy patients.
When needed for therapy, DDAVP is preferred over recombinant vasopressin due to its longer half-life and selective action through the V2 receptor. Selective V2 activity yields little effect on blood pressure and avoids the arterial hypertension encountered with vasopressin administration.
In contrast, there are fewer drugs that are clearly established as being capable of stimulating or inhibiting oxytocin release. Ethanol and methallibure are well known as inhibitors of the release of oxytocin; based on this, ethanol has been used clinically to treat premature labor. While some investigators11 have demonstrated that oxytocin is released when prostaglandin E2 or F2α is given intravenously at term, Dawood et al. report they were unable to stimulate oxytocin release when prostaglandin was given intra-amniotically for the induction of midtrimester abortion (unpublished data).
ATOSIBAN
Preterm labor
If functional oxytocin receptors are crucial to the initiation and propagation of labor, a blockade with a specific receptor antagonist during the last days of pregnancy could be expected to delay and prolong parturition. Several clinical studies85, 86, 87 have examined the efficacy and safety of the oxytocin antagonist I-deamino-2-D-tyr(OET)-4-thr-8-orn-vasotocin/oxytocin (atosiban) for inhibiting preterm labor. This oxytocin antagonist initially appeared promising as a clinical tocolytic because it was able to acutely stop progressive preterm labor in women when compared to other commonly used tocolytics.81, 82, 83 More recently, a Cochrane review failed to demonstrate superiority of atosiban over betamimetics or placebo when six prospective trials of atosiban were analysed.88
Assisted reproductive technology
Atosibsan has also been proposed to antagonize uterine contractions at the time of embryo transfer in assisted reproductive technologies.89, 90 Use of antagonists for oxytocin and vasopressin V1 (V1A) has been reviewed.91 Further investigation is needed before this practice enters standard clinical protocols.
ACKNOWLEDGMENT
The original chapter on the posterior pituitary for the The Global Library of Women’s Medicine was authored by Yusoff M. Dawood, MB ChB, MMed, MD and Firyal S. Khan-Dawood, PhD. They last updated the chapter in 2009. This current presentation is a new editing and update of their original chapter. They continue to deserve credit and my appreciation for the extensive review of these topics they originally prepared and my thanks for the opportunity to provide this update.
REFERENCES
George JM. Immunoreactive vasopressin and oxytocin: concentration in individual human hypothalamic nuclei. Science 1978;200:342-3 |
|
Dierickx K, Vandesande F. Immunocytochemical demonstration of separate vasopressin-neurophysin and oxytocin-neurophysin neurons in the human hypothalamus. Cell and tissue research 1979;196:203-12. |
|
Swaab DF, Nijveldt F, Pool CW. Distribution of oxytocin and vasopressin in the rat supraoptic and paraventricular nucleus. The Journal of endocrinology 1975;67:461-2 |
|
Mohr E, Meyerhof W, Richter D. The hypothalamic hormone oxytocin: from gene expression to signal transduction. Reviews of physiology, biochemistry and pharmacology 1992;121:31-48. |
|
Acher R. Molecular evolution of biologically active polypeptides. Proc R Soc Lond B Biol Sci 1980;210:21-43. |
|
Riddell DC, Mallonee R, Phillips JA, Parks JS, Sexton LA, Hamerton JL. Chromosomal assignment of human sequences encoding arginine vasopressin-neurophysin II and growth hormone releasing factor. Somatic cell and molecular genetics 1985;11:189-95 |
|
Inoue T, Kimura T, Azuma C, et al. Structural organization of the human oxytocin receptor gene. The Journal of biological chemistry 1994;269:32451-6. |
|
Young WS, 3rd, Reynolds K, Shepard EA, Gainer H, Castel M. Cell-specific expression of the rat oxytocin gene in transgenic mice. Journal of neuroendocrinology 1990;2:917-25 |
|
Burford GD, Pickering BT. Intra-axonal transport and turnover of neurophysins in the rat. A proposal for a possible origin of the minor neurophysin component. The Biochemical journal 1973;136:1047-52. |
|
Acher R, Chauvet J, Rouille Y. Dynamic processing of neuropeptides: sequential conformation shaping of neurohypophysial preprohormones during intraneuronal secretory transport. Journal of molecular neuroscience : MN 2002;18:223-8. |
|
Swann RW, Pickering BT. Incorporation of radioactive precursors into the membrane and contents of the neurosecretory granules of the rat neurohypophysis as a method of studying their fate. The Journal of endocrinology 1976;68:95-108 |
|
Zimmerman EA, Carmel PW, Husain MK, et al. Vasopressin and neurophysin: high concentrations in monkey hypophyseal portal blood. Science 1973;182:925-7. |
|
Burbach JP, Adan RA, de Bree FM. Regulation of oxytocin gene expression and forms of oxytocin in the brain. Annals of the New York Academy of Sciences 1992;652:1-13. |
|
Vincent JL, Su F. Physiology and pathophysiology of the vasopressinergic system. Best practice & research Clinical anaesthesiology 2008;22:243-52. |
|
Poole CJ, Carter DA, Vallejo M, Lightman SL. Atrial natriuretic factor inhibits the stimulated in-vivo and in-vitro release of vasopressin and oxytocin in the rat. The Journal of endocrinology 1987;112:97-102. |
|
Mikiten TM, Douglas WW. Effect of calcium and other ions on vasopressin release from rat neurohypophyses stimulated electrically in vitro. Nature 1965;207:302. |
|
Seybold VS, Miller JW, Lewis PR. Investigation of a dopaminergic mechanism for regulating oxytocin release. The Journal of pharmacology and experimental therapeutics 1978;207:605-10. |
|
Verbalis JG, McCann MJ, McHale CM, Stricker EM. Oxytocin secretion in response to cholecystokinin and food: differentiation of nausea from satiety. Science 1986;232:1417-9. |
|
O'Byrne KT, Eltringham L, Clarke G, Summerlee AJ. Effects of porcine relaxin on oxytocin release from the neurohypophysis in the anaesthetized lactating rat. The Journal of endocrinology 1986;109:393-7. |
|
Weitzman RE, Firemark HM, Glatz TH, Fisher DA. Thyrotropin-releasing hormone stimulates release of arginine vasopressin and oxytocin in vivo. Endocrinology 1979;104:904-7. |
|
Dawood MY, Ylikorkala O, Trivedi D, Fuchs F. Oxytocin in maternal circulation and amniotic fluid during pregnancy. The Journal of clinical endocrinology and metabolism 1979;49:429-34. |
|
Gibbens GL, Chard T. Observations on maternal oxytocin release during human labor and the effect of intravenous alcohol administration. American journal of obstetrics and gynecology 1976;126:243-6. |
|
Pawlikowski M, Majak J, Stepien H. Influence of vasopressin and oxytocin upon mitotic activity of adenohypophyseal cells in rat. Endokrynologia Polska 1975;26:417-20. |
|
Dawood MY, Ylikorkala O, Trivedi D, Gupta R. Oxytocin levels and disappearance rate and plasma follicle-stimulating hormone and luteinizing hormone after oxytocin infusion in men. The Journal of clinical endocrinology and metabolism 1980;50:397-400. |
|
Samson WK, Lumpkin MD, McCann SM. Evidence for a physiological role for oxytocin in the control of prolactin secretion. Endocrinology 1986;119:554-60. |
|
Coiro V, Gnudi A, Volpi R, et al. Oxytocin enhances thyrotropin-releasing hormone-induced prolactin release in normal menstruating women. Fertility and sterility 1987;47:565-9. |
|
Suh BY, Liu JH, Rasmussen DD, Gibbs DM, Steinberg J, Yen SS. Role of oxytocin in the modulation of ACTH release in women. Neuroendocrinology 1986;44:309-13. |
|
Barberis C, Mouillac B, Durroux T. Structural bases of vasopressin/oxytocin receptor function. The Journal of endocrinology 1998;156:223-9. |
|
Zeeman GG, Khan-Dawood FS, Dawood MY. Oxytocin and its receptor in pregnancy and parturition: current concepts and clinical implications. Obstetrics and gynecology 1997;89:873-83. |
|
Kimura T, Tanizawa O, Mori K, Brownstein MJ, Okayama H. Structure and expression of a human oxytocin receptor. Nature 1992;356:526-9. |
|
Marc S, Leiber D, Harbon S. Carbachol and oxytocin stimulate the generation of inositol phosphates in the guinea pig myometrium. FEBS letters 1986;201:9-14. |
|
Tom N, Assinder SJ. Oxytocin in health and disease. The international journal of biochemistry & cell biology 2010;42:202-5. |
|
Sabatier N, Rowe I, Leng G. Central release of oxytocin and the ventromedial hypothalamus. Biochemical Society transactions 2007;35:1247-51. |
|
Meyer-Lindenberg A, Domes G, Kirsch P, Heinrichs M. Oxytocin and vasopressin in the human brain: social neuropeptides for translational medicine. Nature reviews Neuroscience 2011;12:524-38. |
|
de Keyzer Y, Auzan C, Lenne F, et al. Cloning and characterization of the human V3 pituitary vasopressin receptor. FEBS letters 1994;356:215-20. |
|
Chard T, Boyd NR, Forsling ML, McNeilly AS, Landon J. The development of a radioimmunoassay for oxytocin: the extraction of oxytocin from plasma, and its measurement during parturition in human and goat blood. The Journal of endocrinology 1970;48:223-34 |
|
Fabian M, Forsling ML, Jones JJ, Pryor JS. The clearance and antidiuretic potency of neurohypophysial hormones in man, and their plasma binding and stability. The Journal of physiology 1969;204:653-68. |
|
Dawood MY, Raghavan KS, Pociask C. Radioimmunoassay of oxytocin. The Journal of endocrinology 1978;76:261-70. |
|
Edwards CR, Chard T, Kitau MJ, Forsling ML, Landon J. The development of a radioimmunoassay for arginine-vasopressin: production of antisera and labelled hormone; separation techniques; specificity and sensitivity of the assay in aqueous solution. The Journal of endocrinology 1972;52:279-88. |
|
Robertson GL, Mahr EA, Athar S, Sinha T. Development and clinical application of a new method for the radioimmunoassay of arginine vasopressin in human plasma. The Journal of clinical investigation 1973;52:2340-52. |
|
Skowsky WR, Rosenbloom AA, Fisher DA. Radioimmunoassay measurement of arginine vasopressin in serum: development and application. The Journal of clinical endocrinology and metabolism 1974;38:278-87. |
|
Soloff MS, Beauregard G, Potier M. Determination of the functional size of oxytocin receptors in plasma membranes from mammary gland and uterine myometrium of the rat by radiation inactivation. Endocrinology 1988;122:1769-72. |
|
Baylis PH, Robertson GL. Plasma vasopressin response to hypertonic saline infusion to assess posterior pituitary function. Journal of the Royal Society of Medicine 1980;73:255-60. |
|
Murphy MR, Seckl JR, Burton S, Checkley SA, Lightman SL. Changes in oxytocin and vasopressin secretion during sexual activity in men. The Journal of clinical endocrinology and metabolism 1987;65:738-41 |
|
Carmichael MS, Humbert R, Dixen J, Palmisano G, Greenleaf W, Davidson JM. Plasma oxytocin increases in the human sexual response. The Journal of clinical endocrinology and metabolism 1987;64:27-31 |
|
Fox CA, Knaggs GS. Milk-ejection activity (oxytocin) in peripheral venous blood in man during lactation and in association with coitus. The Journal of endocrinology 1969;45:145-6. |
|
Levin RJ. Can the controversy about the putative role of the human female orgasm in sperm transport be settled with our current physiological knowledge of coitus? The journal of sexual medicine 2011;8:1566-78. |
|
Miller FD, Chibbar R, Mitchell BF. Synthesis of oxytocin in amnion, chorion and decidua: a potential paracrine role for oxytocin in the onset of human parturition. Regulatory peptides 1993;45:247-51. |
|
Chibbar R, Miller FD, Mitchell BF. Synthesis of oxytocin in amnion, chorion, and decidua may influence the timing of human parturition. The Journal of clinical investigation 1993;91:185-92. |
|
Dawood MY, Wang CF, Gupta R, Fuchs F. Fetal contribution to oxytocin in human labor. Obstetrics and gynecology 1978;52:205-9. |
|
Seppala M, Aho I, Tissari A, Ruoslahti E. Radioimmunoassay of oxytocin in amniotic fluid, fetal urine, and meconium during late pregnancy and delivery. American journal of obstetrics and gynecology 1972;114:788-95 |
|
Kumaresan P, Anandarangam PB, Dianzon W, Vasicka A. Plasma oxytocin levels during human pregnancy and labor as determined by radioimmunoassay. American journal of obstetrics and gynecology 1974;119:215-23. |
|
Coch JA, Brovetto J, Cabot HM, Fielitz CA, Caldeyro-Barcia R. Oxytocin-Equivalent Activity in the Plasma of Women in Labor and during the Puerperium. American journal of obstetrics and gynecology 1965;91:10-7. |
|
Dawood MY, Raghavan KS, Pociask C, Fuchs F. Oxytocin in human pregnancy and parturition. Obstetrics and gynecology 1978;51:138-43. |
|
Noddle BA. Transfer of Oxytocin from the Maternal to the Foetal Circulation in the Ewe. Nature 1964;203:414. |
|
Dawood MY, Lauersen NH, Trivedi D, Ylikorkala O, Fuchs F. Studies of oxytocin in the baboon during pregnancy and delivery. Acta endocrinologica 1979;91:704-18. |
|
Boyd NR, Chard T. Human urine oxytocin levels during pregnancy and labor. American journal of obstetrics and gynecology 1973;115:827-9. |
|
Dicker SE, Tyler C. Vasopressor and oxytocic activities of the pituitary glands of rats, guinea-pigs and cats and of human foetuses. The Journal of physiology 1953;121:206-14. |
|
Khan-Dawood FS, Dawood MY. Oxytocin content of human fetal pituitary glands. American journal of obstetrics and gynecology 1984;148:420-3. |
|
Pavel S. Vasotocin biosynthesis by neurohypophysial cells from human fetuses. Evidence for its ependymal origin. Neuroendocrinology 1975;19:150-9. |
|
Sende P, Pantelakis N, Suzuki K, Bashore R. Plasma oxytocin determinations in pregnancy with diabetes insipidus. Obstetrics and gynecology 1976;48:38S-41S. |
|
Nishimori K, Young LJ, Guo Q, Wang Z, Insel TR, Matzuk MM. Oxytocin is required for nursing but is not essential for parturition or reproductive behavior. Proceedings of the National Academy of Sciences of the United States of America 1996;93:11699-704. |
|
Dawood MY, Khan-Dawood FS, Wahi RS, Fuchs F. Oxytocin release and plasma anterior pituitary and gonadal hormones in women during lactation. The Journal of clinical endocrinology and metabolism 1981;52:678-83. |
|
MacMillan JB, 3rd, Hale RW. Contraction stress testing with mammary self-stimulation. The Journal of reproductive medicine 1984;29:219-21 |
|
Rosenzweig BA, Levy JS, Schipiour P, Blumenthal PD. Comparison of the nipple stimulation and exogenous oxytocin contraction stress tests. A randomized, prospective study. The Journal of reproductive medicine 1989;34:950-4. |
|
Huddleston JF, Sutliff G, Robinson D. Contraction stress test by intermittent nipple stimulation. Obstetrics and gynecology 1984;63:669-73. |
|
Kadar N, Tapp A, Wong A. The influence of nipple stimulation at term on the duration of pregnancy. Journal of perinatology : official journal of the California Perinatal Association 1990;10:164-6. |
|
Stein JL, Bardeguez AD, Verma UL, Tegani N. Nipple stimulation for labor augmentation. The Journal of reproductive medicine 1990;35:710-4 |
|
Di Lieto A, Miranda L, Ardito P, Favale P, Albano G. Changes in the Bishop score induced by manual nipple stimulation. A cross-over randomized study. Clinical and experimental obstetrics & gynecology 1989;16:26-9. |
|
Wathes DC, Swann RW, Pickering BT, Porter DG, Hull MG, Drife JO. Neurohypophysial hormones in the human ovary. Lancet 1982;2:410-2. |
|
Khan-Dawood FS, Dawood MY. Human ovaries contain immunoreactive oxytocin. The Journal of clinical endocrinology and metabolism 1983;57:1129-32. |
|
Dawood MY, Khan-Dawood FS. Human ovarian oxytocin: its source and relationship to steroid hormones. American journal of obstetrics and gynecology 1986;154:756-63. |
|
Ivell R, Furuya K, Brackmann B, Dawood Y, Khan-Dawood F. Expression of the oxytocin and vasopressin genes in human and baboon gonadal tissues. Endocrinology 1990;127:2990-6. |
|
Khan-Dawood FS, Marut EL, Dawood MY. Oxytocin in the corpus luteum of the cynomolgus monkey (Macaca fascicularis). Endocrinology 1984;115:570-4. |
|
Khan-Dawood FS, Huang JC, Dawood MY. Baboon corpus luteum oxytocin: an intragonadal peptide modulator of luteal function. American journal of obstetrics and gynecology 1988;158:882-91. |
|
Khan-Dawood FS, Goldsmith LT, Weiss G, Dawood MY. Human corpus luteum secretion of relaxin, oxytocin, and progesterone. The Journal of clinical endocrinology and metabolism 1989;68:627-31. |
|
Auletta FJ, Paradis DK, Wesley M, Duby RT. Oxytocin is luteolytic in the rhesus monkey (Macaca mulatta). Journal of reproduction and fertility 1984;72:401-6. |
|
Ang VT, Jenkins JS. Neurohypophysial hormones in the adrenal medulla. The Journal of clinical endocrinology and metabolism 1984;58:688-91. |
|
Sharma OP, Hays RL. A possible role for oxytocin in sperm transport in the male rabbit. The Journal of endocrinology 1976;68:43-7. |
|
Adashi EY, Hsueh AJ. Direct inhibition of testicular androgen biosynthesis revealing antigonadal activity of neurohypophysial hormones. Nature 1981;293:650-2. |
|
Legros JJ. The radioimmunoassay of neurophysins as a tool for the posterior pituitary investigation. Biochemical pharmacology 1978;27:1009-18. |
|
Robinson AG, Archer DF, Tolstoi LF. Neurophysin in women during oxytocin-related events. The Journal of clinical endocrinology and metabolism 1973;37:645-52. |
|
Khan-Dawood FS. Immunocytochemical localization of oxytocin and neurophysin in human corpora lutea. The American journal of anatomy 1987;179:18-24. |
|
Khan-Dawood FS. Localization of oxytocin and neurophysin in baboon (Papio anubis) corpus luteum by immunocytochemistry. Acta endocrinologica 1986;113:570-5. |
|
Akerlund M, Stromberg P, Hauksson A, et al. Inhibition of uterine contractions of premature labour with an oxytocin analogue. Results from a pilot study. British journal of obstetrics and gynaecology 1987;94:1040-4. |
|
Andersen LF, Lyndrup J, Akerlund M, Melin P. Oxytocin receptor blockade: a new principle in the treatment of preterm labor? American journal of perinatology 1989;6:196-9. |
|
Goodwin TM, Paul R, Silver H, et al. The effect of the oxytocin antagonist atosiban on preterm uterine activity in the human. American journal of obstetrics and gynecology 1994;170:474-8. |
|
Papatsonis D, Flenady V, Cole S, Liley H. Oxytocin receptor antagonists for inhibiting preterm labour. Cochrane Database Syst Rev 2005:CD004452. |
|
Pierzynski P, Reinheimer TM, Kuczynski W. Oxytocin antagonists may improve infertility treatment. Fertility and sterility 2007;88:213 e19-22. |
|
Moraloglu O, Tonguc E, Var T, Zeyrek T, Batioglu S. Treatment with oxytocin antagonists before embryo transfer may increase implantation rates after IVF. Reproductive biomedicine online 2010;21:338-43. |
|
Pierzynski P. Oxytocin and vasopressin V(1A) receptors as new therapeutic targets in assisted reproduction. Reproductive biomedicine online 2011;22:9-16. |