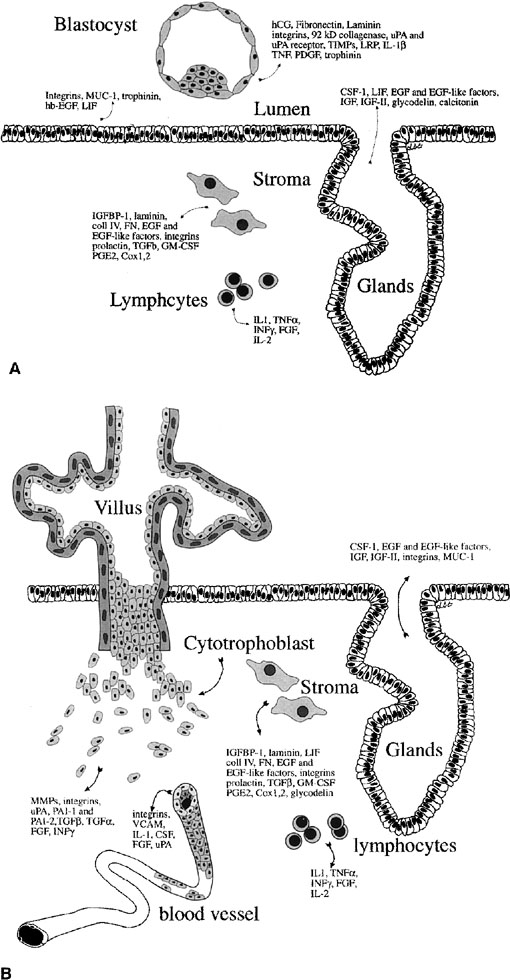
Implantation
Authors
INTRODUCTION
The attachment and implantation of an embryo into a mother’s uterus is the critical event that defines an embryo’s fate. Implantation is not an efficient process, and is successful only about 10–15% of the time.1 As a hallmark of development, implantation marks the very beginning of life, and proceeds from the tentative first attachment reaction (apposition) to subsequent aggressive embryonic intrusion and invasion into the maternal decidua. The earliest events of implantation have been difficult to study because the very act of examination disrupts the nature of the process. Animal models have shown a remarkable dissimilarity in implantation strategies, even when relatively closely related species are compared. However, with the development of modern molecular biology techniques and with the use of the donor egg model from assisted reproductive technologies (ART), we are now gaining increasing knowledge about implantation, especially at the biochemical level. We now know that synchronized development of both endometrium and embryo is a prerequisite for implantation to succeed, and that disruption of this developmental synchrony can result in infertility and/or pregnancy loss.
The events of early pregnancy are tightly coordinated by means of endocrine, paracrine, juxtacrine, and autocrine cellular communication pathways. Adhesion molecules and ligands promote cell-cell attachment and invasion, while proteases digest extracellular matrix (ECM) ahead of the invading embryo. The invading cells of the embryo, called trophoblasts, invade in a fashion not unlike cancer cells,2 however, because of the extensive signaling mechanisms that exist between maternal and embryonic cells, uncontrolled spread and immunologic rejection are avoided. In this chapter we briefly describe a variety of animal models of implantation, then outline the classic studies that form the foundation of our current understanding of human implantation. Finally, we detail the known biochemical markers of uterine receptivity and describe their regulation. Our aim is to maintain this information within a context of clinical applicability for current treatment of infertility and recurrent pregnancy loss and to explore potential future directions of implantation-related research.
ANIMAL MODELS OF IMPLANTATION
Historically, animal models have been the primary means for investigating implantation and they provide insight into various aspects of the process.3, 4, 5 Murid rodents can be induced to exhibit delayed implantation as a result of changes in hormonally controlled uterine development. This has been exploited as a model to study embryo-endometrial communication.6, 7 Implantation in mice occurs on the fourth day after coitus coincident with a rise in serum estrogen levels. Many of the proteins involved in human implantation were first identified in the mouse.8 The mouse is also an excellent model for the study of apoptosis during implantation because epithelial cells undergo programmed cell death after embryo apposition to allow penetration of the trophoblasts into the uterine basal lamina.9
Guinea pigs exhibit a different type of implantation, with similarities to the human. Here the syncytiotrophoblasts put forth appendages that extend through the zona pellucida to achieve apposition. Once contact is made, these appendages invade between uterine epithelial cells into the basal lamina and underlying stroma, thus making the guinea pig a useful model for the study of epithelial penetration.10
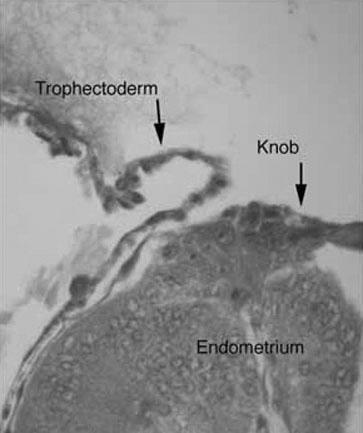
The rabbit has served as an outstanding model to study implantation. As induced ovulators (ovulating approximately 10 hours after mating), implantation in rabbits can be timed reliably.11 Rabbit blastocysts are exceptionally large and form knob-like attachment sites with the uterine epithelium (Fig. 1).12 Such specialized areas of attachment allow for the study of the cell-cell adhesion molecules and their regulation.
{kind=link}
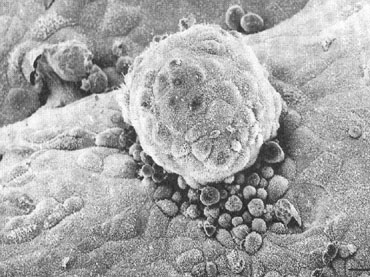
There have also been an increasing number of in vitro models to study implantation using purified endometrial epithelium and cytotrophoblast, but these have had limitations in their use.13, 14, 15 Such models may provide the most intimate look at the earliest moments of embryo-endometrial interaction (Fig. 2). Through the collective use of these various models, much has been learned about the basic principles of implantation. We discuss next some of the classic studies that established the timing of implantation as well as the stages of implantation in humans.
{kind=link}
TIMING AND STAGES OF IMPLANTATION
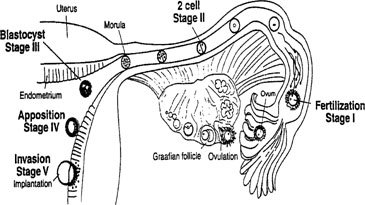
Our current understanding of human implantation has evolved from the pioneering work of Hertig and colleagues 50 years ago, to the present day receptor-mediated model with a more defined estimate of the endometrial window of implantation as described recently by Yoshinaga and Wilcox, respectively.16, 17, 18 Based on work from the infertility and ART clinics, we now know that embryo development is also critical before the overall window of receptivity is achieved.19 The sequence of events from the release of an ovum to the point of established pregnancy has been divided into discreet stages of implantation. As shown in Figure 3, these stages consist of preimplantation events for both the embryo and the endometrium, followed by apposition and attachment, and finally penetration.
{kind=link}
Preimplantation events of the embryo
The earliest aspects of implantation in the human encompass the sequence of events that lead from fertilization to blastocyst attachment and invasion. After ovulation, the nascent oocyte is transported through the Fallopian tube, where fertilization occurs, thus defining stage I of implantation. Stage II is marked by the subsequent initiation of cell division in the embryo. At stage III, the ball of embryonic cells, now called a morula, enters the uterine cavity where further divisions result in formation of the blastocyst. It is estimated that only 20% of human embryos ever reach this point of development but for those that do, activation of the genome begins that is essential for successful implantation.20, 21 These initial events occur within a narrow time frame, as the embryo enters the uterine cavity at 72–96 hours after fertilization.20, 22
Once in the uterine cavity, the embryo remains free for roughly 3 days before attaching to the endometrium.20 During this time the embryo begins its communication with the mother even prior to attachment to the endometrium. Maternal secretions likely nurture the growing embryo.23, 24, 25 Human chorionic gonadotropin (hCG), produced by the embryo even before hatching from the zona pellucida (ZP), is the pregnancy recognition signal to the mother.26, 27 The ZP is important in prevention of polyspermy as well as providing an appropriate environment for early embryonic development; ultimately the embryo must hatch from the ZP prior to apposition, exposing surface receptors and ligands that are important for attachment. The process of hatching becomes less efficient with advancing maternal age and it is thought that zona hardening can entrap an embryo within the ZP.28, 29, 30 Several methods of assisted hatching techniques have been suggested to alleviate this problem in in vitro fertilization (IVF). Such methods to weaken the ZP allow for zona shedding that has improved implantation rates, in particular for older women or for prior implantation failures.
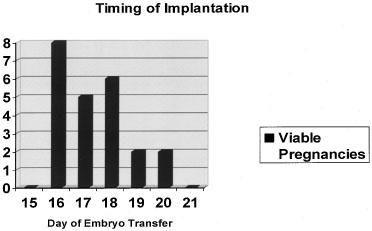
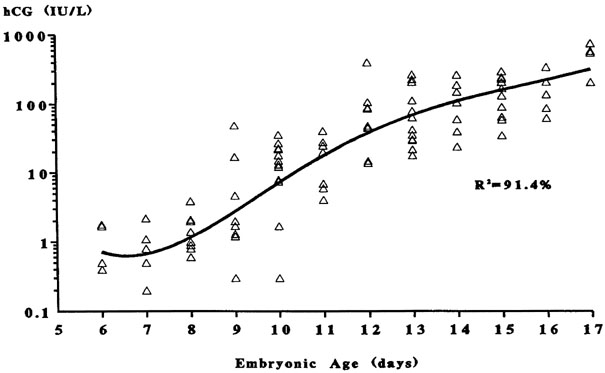
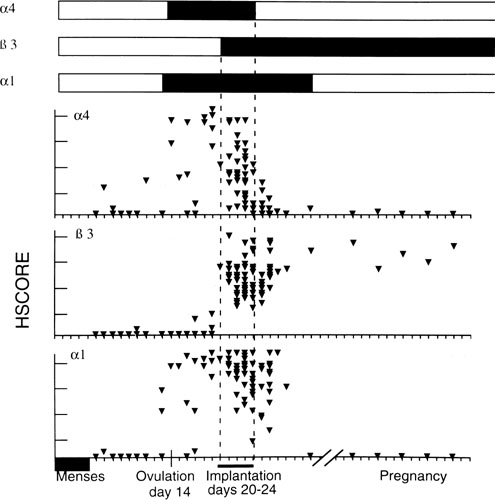
This progression of preimplantation events in the embryo is essential for implantation, and occurs within a narrow time frame during the midsecretory phase. As addressed in later sections, the endometrium is now thought to become permissive to implantation during cycle days 20–24 of a normalized 28-day cycle, the so-called window of receptivity.18 Navot and colleagues31 demonstrated that pregnancies can be achieved by embryo transfers (ET) in donor egg cycles on cycle days 15–20, with the vast majority of success between days 16 and 19 (Fig. 4).31 Actual implantation takes place between embryonic days 5 and 6, so the transfer of 2–3-day-old embryos prior to day 20 yields peak implantation rates.31 This concept was further expanded by this group by studying the timing the first hCG detection in donor egg recipients.19 They found the average detection of pregnancy on embryonic day 7 with 2-day-old embryos transferred on day 15 having hCG detection on day 20 and 2-day-old embryos transferred on day 19 having hCG detection on days 24.19 No detection was found when embryos were transferred after day 19. These data correlate well with a window of receptivity between cycle days 20–24 and agree with the studies by Wilcox and colleagues18 who also studied the timing of implantation. Such donor-recipient models are able to establish the triphasic pattern of early hCG, rise which has clinical applicability for many gynecologic problems such as ectopic pregnancies, abnormal intrauterine pregnancies, and gestational trophoblastic disease (Fig. 5).19
{kind=link}
{kind=link}
Preimplantation events of the uterus
STRUCTURAL CHANGES OF THE ENDOMETRIUM
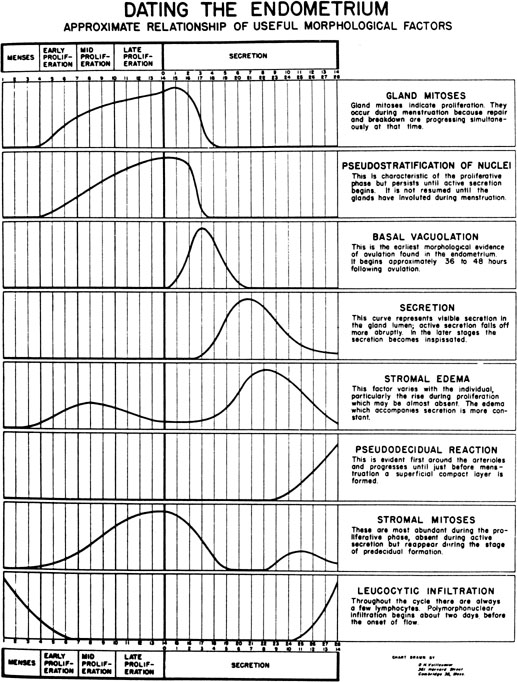
During the first half of the menstrual cycle, called the proliferative phase, the endometrium thickens in response to rising estradiol levels (Fig. 6). As assessed by transvaginal ultrasound, the endometrium grows rapidly during the first 14 days of the cycle. It is thought that an endometrial thickness less than 6 mm is unlikely to support a pregnancy.32, 33 Once ovulation takes place and the granulosa transforms into the corpus luteum (CL), the endometrium is converted to a secretory structure under the influence of rising progesterone. This conversion follows an orderly progression as demonstrated by Noyes and coworkers34 in their now classic paper on endometrial dating (Fig. 7). Initially, the endometrial glandular epithelium begins to synthesize and secrete glycogen and glycoproteins, and by midcycle, stromal edema develops, accompanied by proliferation of the spiral arterioles. Ultrastructurally, scanning electron microscopy of the midluteal luminal epithelium shows the development of small protrusions called pinopods, the appearance of which coincides well with the suspected time of maximal receptivity (Fig. 8). Such surface modifications are thought to play a functional role in embryo apposition and attachment.35, 36, 37
{kind=link}
{kind=link}
{kind=link}
Another progesterone-related uterine change is the frequency of uterine contractions. Animal studies have shown decreased implantation rates with increasing uterine contraction.38, 39 Work by Fanchin and coworkers40 using digitized 5-minute ultrasound scans showed that contractility is inversely related to progesterone levels in humans as well. They also showed decreased clinical pregnancy rates with increasing uterine contractions. Lesny and coworkers41 also studied this phenomenon doing mock ET using 30 μL of an echogenic fluid medium. They noted that with easy transfers, uterine contractions remained at a stable baseline and the medium did not migrate significantly. However, difficult transfers were associated with strong, random contractions and the fluid medium migrated from the fundal region in six of seven patients (in two the medium migrated to the Fallopian tubes).41 This is still a relatively new concept but may lead to future therapeutic considerations in ART.
BIOCHEMICAL CHANGES OF THE ENDOMETRIUM
Aside from what is known about the structural changes that the endometrium undergoes during the secretory phase, there is also a growing body of information regarding the biochemical changes of the endometrium. Discoveries regarding the ECM, cell adhesion molecules, cytokines and growth factors, and the enzymes that degrade the ECM have shed light on embryo-endometrial interactions that occur during attachment and invasion.
Extracellular matrix
The ECM is formed from secreted proteins and glycoproteins, and forms the ground substance outside the cells in all tissues. The ECM appears to play an important role in the cell-cell interactions.42 Analysis of these substances in the endometrium throughout the menstrual cycle has shown constitutive expression of collagen types III and V, and fibronectin.42 Cycle-dependent variations in expression also exist; type VI collagen is produced throughout the proliferative phase, but declines during the secretory phase. There may also be differences in the expression of these various ECM components in endometrium of fertile and infertile women.43 The luminal surface is an effective barrier to implantation during much of the reproductive cycle.39 Early on, it was noticed that there is thinning of the carbohydrate-rich domain of the luminal uterine surface, known as the glycocalyx, and a decreased negative charge at the luminal epithelium at the midluteal phase.44, 45 Variations in the ECM that make up these phenomena may hinder embryo-epithelial contact necessary for apposition. One mucin component in particular, MUC-1, may serve as an antiadhesive molecule and limit apposition.46 In rodents, MUC-1 disappears at the time of implantation and persistent expression results in implantation failure. Studies in human endometrium have been less convincing because MUC-1 persists throughout the luteal phase in women. Nevertheless, there may still be a role of MUC-1 during human implantation.47
Matrix metalloproteinases and their inhibitors
Matrix metalloproteinases (MMPs) are zinc-containing endopeptidases that enzymatically digest certain ECM proteins and therefore play an important role in tissue remodeling processes.48, 49 There are at least three groups of these enzymes differentiated by their substrates, as well as a group of membrane-type matrix metalloproteinases (MT-MMPs) (Table 1). Endometrial expression of MMPs is highly regulated to facilitate successful implantation or timely degradation and remodeling of the endometrial lining during menstruation, in the absence of a successful pregnancy.50, 51
Table 1. Matrix metalloproteinases
Class | Name | Substrate | Cell Source |
Collagenases | MMP-1 | Collagen I, II, III, VII, VIII, and X | Endometrial stroma, cytotrophoblast |
MMP-8 | Same as MMP-1 | Same as MMP-1 | |
Gelatinases | MMP-2 (gelatinase A) | Collagen IV gelatin | Endometrial stroma, cytotrophoblast |
MMP-9 (gelatinase B) | Same as MMP-2 | Same as MMP-2 | |
Stromelysin | MMP-3 (stromelysin-1) | Fibronectins, laminins, collagens III, IV, V, elastin, proteoglycan | Endometrial stroma |
MMP-10 (stromelysin-2) | Same as MMP-3 | Same as MMP-3 | |
MMP-11 (stromelysin-3) | ? | Endometrial epithelium | |
MMP-7 (matrilysin) | Collagen IV, gelatin, fibronectin, proteoglycan | Proliferative endometrial epithelium | |
Membrane-type MMPs | MT-MMP-1 | ? | |
MT-MMP-2 | |||
MT-MMP-3 |
MMP, matrix metalloproteinases.
MMPs are produced as inactive precursors that require activation by other peptidases, denaturants, or heat.52 This cascade of activation is often initiated by the MT-MMPs and is tightly controlled by a variety of tissue inhibitors of metalloproteinases (TIMPs).53 The ECM constituents can actually regulate cellular activity through a receptor-mediated process, thus, production and breakdown of the ECM influences a tissue’s characteristics through a dynamic reciprocity and is vital to invasion and maternal recognition of a pregnancy.50, 54 There are also other interactions between the MMPs and the embryo-endometrial complex that are likely involved with implantation. Cell adhesion receptors known as integrins appear to have a role in MMP expression and function. Engagement of a placental integrin with fibronectin induces MMP expression and the integrin αvβ3 directly binds to and activates MMP-2.55, 56 MMP-2 is one of the most active MMPs in the distal invasive cell column of human trophoblasts during implantation and allows these cells to invade in the direction of their migration.57 MMP-9 is also essential for both rodent and human placental invasion.58 Decidual cells strongly express the MMP inhibitor TIMP-2, presumably as part of the maternal mechanism to block uncontrolled invasion of the placenta.59
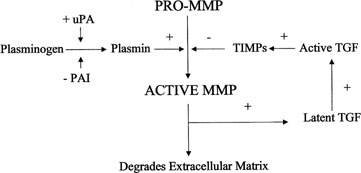
The delicate interplay between these factors must be precisely coordinated for successful but controlled invasion of the trophoblast. Plasminogen, its activators, and its inhibitors appear to participate in this process.60 Plasmin, a protease derived from plasminogen, activates certain MMPs and has a role in degradation of the ECM. Regulation of plasminogen to plasmin conversion is stimulated by urokinase-type plasminogen activator (uPA) and inhibited by alpha2-macroglobulin (α2MG). Human endometrium, embryos, and trophoblasts express both uPA and its receptor, and mutations in these have been associated with implantation failure.61, 62, 63, 64, 65 Activity of plasminogen activator (PA) is tightly regulated by specific inhibitors of PAs (PAI-1 and PAI-2), which bind and inactivate PAs.66 Plasmin activates transforming growth factor β (TGF-β), which in turn activates TIMP and PAI production, thus inhibiting further plasmin activation (Fig. 9). Polycystic ovary syndrome (PCOS) is associated with elevated PAI-1 activity, which may contribute to the higher than expected rate of pregnancy loss via this pathway.67, 68
{kind=link}
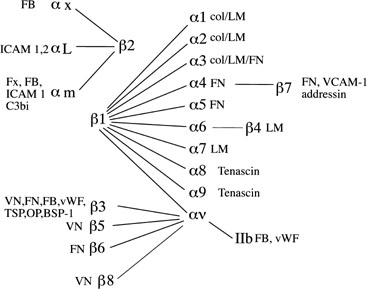
Integrin cell adhesion molecules
Integrins, one member of a larger family of cell adhesion molecules, have been well studied at the levels of the embryo, trophoblast, and endometrial epithelium and stroma throughout the menstrual cycle and into pregnancy (Fig. 10).69, 70, 71, 72, 73, 74, 75, 76 While many integrins are constitutively expressed in human endometrium, the expression of certain integrins coincides well with the period of peak uterine receptivity.71, 72 The timing of receptivity was first described in the classic studies by Hertig and colleagues.16 Over a 15-year time period, they examined 210 hysterectomy specimens, ranging from the day of ovulation to 17 days postovulation. They observed that in uteri obtained prior to cycle day 20, all embryos were free-floating in the tubes or uterus, while in uteri obtained after day 21 the embryos were attached to the uterine lining. Many subsequent studies have helped establish that peak uterine receptivity is between cycle days 20 and 24 of a normalized 28-day cycle.18, 31
{kind=link}
Three integrins (α1β1, α4β1, and αvβ3) have been shown to be coexpressed during the window of receptivity in humans, present together from cycle day 20–24 (Fig. 11). Of these three, the luminal epithelium expresses only αvβ3 at the apical pole, suggesting a potential role in embryo attachment.77 This integrin recognizes the three amino acid sequence arg-gly-asp (RDG), which has been implicated in trophoblast attachment and outgrowth in several studies.78, 79 Interestingly, we have shown a loss of midluteal αvβ3 expression in certain women with unexplained infertility as well as in women with conditions known to have diminished implantation rates, including PCOS, endometriosis, and those with tubal disease and hydrosalpinges, further implicating the critical role of this integrin during implantation.74, 75, 76, 80 Animal studies have also demonstrated that blockade of this integrin will prevent or reduce implantation. This could prove to be useful for future diagnostic and therapeutic considerations for the infertile couple.
{kind=link}
Other markers of uterine receptivity
A growing number of proteins involved in endometrial-embryo interaction both during and after implantation have been identified (Fig. 12).81 As mentioned earlier, mucins like MUC-1 have been suggested as markers of receptivity. Glycodelin (placental protein 14 [PP14]) is a glycoprotein produced by the glandular epithelium and has been associated with inadequate hormonal cycles.82 There are also other adhesion molecules that undergo upregulation during the time of uterine receptivity, such as trophinin, cadherin-11,83, 84 and the hyaluronate receptor, CD44.85 Many cytokines and growth factors, such as leukemia-inhibitory factor (LIF), insulin-like growth factor (IGF)-II, heparin-binding epidermal growth factor (HB-EGF), and interleukin (IL)-1β also appear to be expressed during peak uterine receptivity.86, 87, 88, 89
{kind=link}
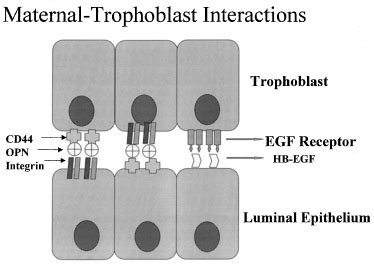
One of the most recently defined markers is a secreted glycoprotein called osteopontin (OPN). This RGD-containing protein has binding sites to two major receptors that are present on both the material and embryonic epithelium, αvβ3 and CD44. Osteopontin is a 70-kd glycosylated phosphoprotein secreted by the glandular epithelium and is expressed during the midsecretory phase, localized to the luminal endometrial epithelium.90 Regulation of OPN is tied to progesterone.90, 91, 92 The apical localization of OPN suggests a role as a sandwich ligand that serves as a bridge binding surface receptors on the endometrial and embryonal surfaces (Fig. 13).
{kind=link}
Apposition
Despite this wide array of structural and biochemical changes by the preimplantation embryo and endometrium, it has been difficult to ascribe specific function to many of them. Apposition, defined as stage IV of implantation, is still an elusive process to observe in humans, and there remains much uncertainty about its mechanism and exact timing. It appears many of the factors described above have critical roles in bringing the endometrium and embryo together. Both express a variety of adhesion molecules and ligands during the expected time of implantation. Current theories suggest that endometrial surface projections, known as pinopods, form a privileged site of receptor expression, raising the endometrial apical surface above anti-adhesive molecules such as MUC-1. Once contact is made between the embryo and the maternal surface, polar trophoblast displaces the endometrial cells by sending ectoplasmic protrusions between them and disrupting their desmosomes.93, 94 This invasiveness appears dependent on the formation of the syncytiotrophoblast, occurring less than 24 hours after embryo adhesion.95
In the vasculature, leukocytes bind to endothelial cells through initial interaction with carbohydrate-binding proteins called selectins. This first step initiates further cellular interactions by integrins and such as are addressed above.96 It has now been demonstrated that human endometrium up-regulates L-selectin during the window of receptivity and is also expressed on human trophoblasts.97 This marker has shown decreased expression in controlled ovarian hyperstimulation and may provide another marker of endometrial function and receptivity.98
Penetration and invasion
Apposition is followed quickly by epithelial penetration and invasion (stage V). Again, this is a process that is not clearly understood but must be tightly orchestrated by a multitude of components previously mentioned, namely the ECM, cell adhesion molecules, MMPs and their inhibitors, and a variety of growth factors and their receptors. The importance of the ECM on cell surface proteolysis has been studied and suggests that on invasion, cell migration and the acquisition of an invasive phenotype may be stimulated by exposed ECM and digested fragments of the endometrial or trophoblast ECM mediated in large part by activation of specific MMPs.99, 100 Rather than a destructive process, invasion of the placental cells appears to involve breaking the intercellular connections and selective apoptosis. The intruding trophoblast appears to adhere to the lateral surfaces of the luminal epithelium with formation of junctional complexes and pushes these cells aside as the mass of the embryo migrates into the underlying decidua.
With time, trophoblast invasion reaches the maternal circulation. Access to the maternal vasculature becomes a priority for the growing embryo, which requires increasing quantities of nutrients and oxygen and better management of cellular waste for its survival. This stage of implantation is marked by rapid expansion of both cytotrophoblast and syncytial trophoblast (Fig. 14).101 At stage Va of invasion, the maternal vasculature remains intact, but becomes surrounded by the expanding syncytium. With further growth, the syncytium and cytotrophoblast invade the maternal vasculature, and the cytotrophoblast is incorporated into the wall of maternal vessels. As detailed later, this ability to mimic endothelial cell characteristics is critical to this invasion, thus establishing a blood supply and a presence within the maternal tissues that will remain intact for the remainder of the pregnancy.
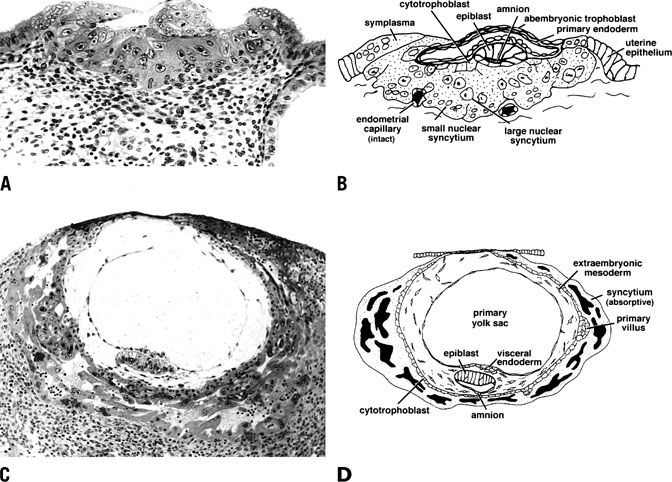{kind=link}
As shown in Figure 14C and 14D, stage Vb of invasion is characterized by expansion of the syncytium and cytotrophoblast and establishment of lacunae as a result of vasculature invasion. By stage Vc, the embryo is fully below the luminal surface and the circumference of the embryo is surrounded by a layer of cytotrophoblast that will rapidly bud to form villi. Development of the placenta and stage V ends by day 11–12 after ovulation with the development of primary villi.
REGULATION OF UTERINE RECEPTIVITY
As discussed, the implantation process is tightly regulated to allow proper attachment and penetration of the embryo while preventing uncontrolled invasion. Ovarian steroids provide the overriding endocrinologic control throughout the menstrual cycle while cytokines and growth factors work at the cellular level as paracrine and autocrine mediators of steroid action. As outlined below, highly coordinated events, involving receptors, cell adhesion molecules, and ECM components coupled with the enzymes and their inhibitors that degrade ECM, regulate implantation.
Steroid hormones and uterine receptivity
Ultimately, implantation of the nascent embryo is dependent on ovarian production of the sex steroids produced by the developing follicle and by the subsequent corpus luteum.102 Estradiol (E2) derived from the developing follicle initiates endometrial growth and cell division increasing epithelial, stromal, and vascular components of the endometrium. Through hypertrophy and hyperplasia the endometrium grows in response to estrogen. After ovulation, progesterone, which is an essential hormone for successful pregnancy, transforms the endometrium into a secretory structure capable of supporting early pregnancy.103
Steroid hormone receptors are expressed by the growing endometrium and dictate the action of the steroid hormones. Estrogen receptors-α and -β (ERα and ERβ), progesterone receptors A and B (PR-A and PR-B), androgen receptors (AR), and glucocorticoid receptors (GR) are all expressed in human endometrium.104, 105, 106 It is primarily ERα and the PRs that play the major role in endometrial development.107, 108 The receptors for estrogen and progesterone are regulated by these hormones in the endometrium. Estrogen upregulates both ERα and the PR with increased concentration of both during the proliferative phase in both epithelial and stromal cells.109, 110 Both ERα and PR are downregulated by progesterone and decline during in the secretory phase. Interestingly, ERα is decreased at the time of implantation in every mammal studied to date and may be a critical event for the establishment of uterine receptivity.111 Although PR has been reported to also be reduced at the time of implantation,112 Mote and colleagues105 recently suggested that it is specifically epithelial PR-A that declines at the time of implantation. Thus, the relative increased activity of ERβ and PR-B by the midsecretory phase may have a role in regulating key implantation-specific genes.104, 105, 113 A diminished role for ERα is consistent with the previous demonstration by DeZiegler and coworkers110 that estrogen is unnecessary for the development of the secretory endometrium. In fact, failure to downregulate ERα may actually be detrimental to implantation, because it has been shown that estrogen inhibits the expression of several endometrial factors, including the integrin β3 subunit and OPN.114
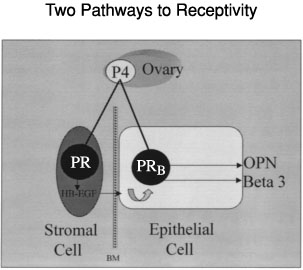
The expression of certain proteins involved in the establishment of uterine receptivity may be regulated through indirect (paracrine) pathways. While an endocrine pathway represents a direct effect of hormones such as progesterone on the estrogen-primed endometrial epithelium, paracrine pathways also can be demonstrated, and represent an indirect effect of progesterone acting through endometrial stroma (Fig. 15). Cunha and colleagues115, 116, 117 demonstrated the importance of endometrial stroma in directing epithelial growth and differentiation using ER and PR knockout mice. In the human endometrium, we have demonstrated that both endocrine and paracrine pathways are functional.118 We have also defined defects in each of these two pathways that may contribute to infertility. Type I defects, as we have called them,72 are a result of deficient ovarian estrogen and/or progesterone and are associated with significant histologic delay as defined by Noyes and colleagues34 (see Fig. 7). In this case progesterone-induced genes such as calcitonin or osteopontin are delayed in their expression. Type II defects represent a separate effect resulting from inappropriate paracrine activity. While endometrial histology appears unaffected, differentiation of the epithelium fails to occur perhaps as a result of an alteration in the action of stromal factors on epithelial cells. An example includes the loss of the αvβ3 integrin seen in some women with infertility and endometriosis.75 These pathways can be independently disrupted but are functionally related. The topic of luteal phase defect (LPD), which is a common cause of both infertility and recurrent pregnancy loss, is the result of these related pathways, and will be expanded in detail in a later section.119
{kind=link}
Cytokines and growth factors
Cytokines and growth factors are peptides that recognize and bind specific cell membrane receptors to signal intracellular events. Many cytokines have been recognized as important during implantation in preimplantation embryos and uteri.120 These factors are important to an increasing list of processes including cellular growth, differentiation, and immunomodulation. While the list of biologic signals in the endometrium continues to grow, several factors have been established as essential for normal implantation.
COLONY STIMULATING FACTOR-1
Colony stimulating factor-1 (CSF-1), initially described as a hematopoietic growth factor, is a 50–70-kDa glycoprotein expressed by fibroblasts, monocytes, macrophages, and endothelial cells.121 The role of CSF-1 in fertility was first described in a mouse model in which mice deficient in CSF-1 were infertile as a result of implantation failure.122 Subsequent human studies have shown that both CSF-1 and its receptor (the c-fms proto-oncogene) are upregulated in the late secretory phase and in early pregnancy; other studies connect CSF-1 to trophoblastic growth and differentiation.123, 124 Thus, the temporal expression of CSF-1 and its receptor, its effect on trophoblasts, and data from CSF-1 null mice suggest an important role for CSF-1 in implantation.
LEUKEMIA-INHIBITORY FACTOR
LIF was originally shown to induce differentiation of a myeloid leukemia cell line.125 Its association with fertility was established when it was discovered that peak uterine expression of LIF occurred at the time of implantation in mice, and in the delayed implantation model, injection of estradiol correlated with an increase in uterine LIF expression.126 Interestingly, while LIF-null mutant male mice are fertile, female mice lacking LIF fail to implant, although their embryos transferred into wild-type mice will result in pregnancy, demonstrating the importance of LIF in the implantation process.86
Human studies have also shown peak uterine expression during the window of implantation that may be decreased in some women with infertility.127 LIF is produced in the luminal epithelium and causes increased trophoblastic production of fetal fibronectin and other proteins, thus implicating it as a direct maternal modulator of trophoblastic differentiation.127, 128
INTERLEUKIN-1 FAMILY
IL-1 represents a family of peptides composed of IL-1α (159 amino acids), IL-1β (153 amino acids), IL-1 receptor antagonist (IL-1ra), and two IL-1 receptors (IL-1R, type I and II).129 IL-1α increases fetal fibronectin production and immunoreactivity of MMP-9 by purified first trimester cytotrophoblasts in vitro.130 IL-1β has been identified in oocytes, embryos, and endometrial stroma, and, along with IL-1ra and IL-1R type I, has been localized to macrophages in the villus trophoblast and maternal-trophoblast interface.131, 132, 133 Both IL-1Rs are found on the luminal epithelium and this could be a pathway through which an embryo can actually facilitate its own implantation. De los Santos and coworkers134 demonstrated that not all embryos express these factors and suggested that embryonic competency relates to the presence or absence of these specific cytokine signals.
Further understanding of the role of IL-1 in implantation has come from the IL-1 receptor antagonist. This peptide is found in large amounts during the follicular phase of the menstrual cycle, but diminishes near the time of implantation.135 Implantation failure occurs in mice injected with exogenous IL-1ra, thus implicating a vital role of IL-1 for successful implantation.136 Interestingly, IL-1R type I null mutant mice display normal implantation, even in the presence of exogenous IL-1ra.137 This may be caused by developmental compensation that overcomes the lack of IL-1 signaling in the null-mutant mice. Alternatively such studies may be demonstrating that IL-1 does not have an essential role during implantation. A precise role of this cytokine during implantation awaits further investigation.
GROWTH FACTORS
Epidermal growth factor and EGF-like molecules
EGF is a 6-kDa peptide that is a potent stimulator of proliferation in a variety of cell types.138 The EGF family of growth factors has expanded to include EGF itself, TGF-α, HB-EGF, amphiregulin, and betacellulen. These factors act through binding to the ErbB receptors (ErbB 1 through 4), each specific for one of the aforementioned ligands.139
The importance of EGF and EGF-like molecules in implantation has been well-demonstrated in the mouse model. It appears that these molecules stimulate embryo development, in that both EGF and TGF-α improve ZP hatching as well as stimulate trophoblast outgrowth in vitro.140, 141 In mice, uterine EGF expression is regulated by estrogen and may be part of the link between the estrogen rise and blastocyst activation/implantation in the delayed implantation model.142 This is further supported by the observation that exogenous EGF can trigger implantation in a similar fashion to exogenous estrogen in this model.143 Inappropriate uterine TGF-α expression in transgenic mice also results in aberrant implantation.144
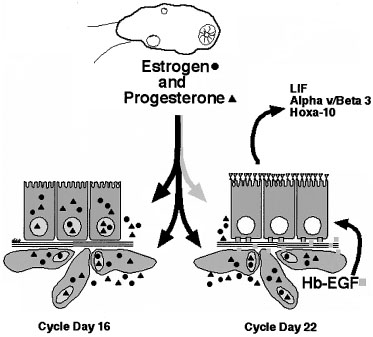
The role of EGF and the EGF-like molecules are less clearly understood during human or primate implantation. Multiple studies show conflicting results regarding timing and location of expression of many of these factors.145, 146, 147 HB-EGF has been shown to be expressed during the window of implantation in multiple species, including human.148 HB-EGF is regulated differentially in the endometrial epithelium and stroma by estrogen and progesterone, respectively.149 Recent data show that HB-EGF is upregulated in endometrial stroma in the midluteal phase and that this stimulates epithelial expression of various important biomarkers of receptivity such as the αvβ3 integrin, HOXA-10, and LIF (Fig. 16).118
{kind=link}
Insulin-like growth factors
The IGF family of growth factors includes two distinct hormones (IGF-I and IGF-II), two IGF receptors, and six insulin-like growth factor binding proteins (IGFBPs). IGF-I and IGF-II are mitogenic growth factors that are structurally similar to insulin and they are present in human endometrium throughout the menstrual cycle, promoting endometrial cellular mitosis and differentiation.150 Both hormones are present in the endometrial stroma, but the receptors are found in both the stroma and epithelium.151 IGF-I is abundant during the proliferative phase and may promote proliferation; conversely, IGF-II is expressed more during the secretory phase.
The IGFBPs bind to and modulate the activity of IGF-I and IGF-II.152 All six are present in human endometrium with IGFBP-1 (also known as PP12 or α1-PEG) being a major secretory protein of the stroma.153 IGFBP-1 is regulated by progesterone and it acts via two mechanisms.154 First, as IGF-I and IGF-II inhibit decidualization, the presence of IGFBP-1 probably counteracts the effect of trophoblast IGF-II. Second, IGFBP-1 contains the integrin-binding domain arg-gly-asp (RGD) and is recognized specifically by the α5β1 integrin present on cytotrophoblast cells and endometrial stroma and decidua.155 These interactions may act to limit invasion of the cytotrophoblast, although much is still unknown about these mechanisms.
Transforming growth factor β
The TGF-β superfamily is a group of related secretory proteins that regulate a variety of cellular processes. This group contains the four TGF-β isoforms (TGF-β1 through TGF-β4), inhibin, activin, Mullerian-inhibiting substance, VEGF-1, endometrial bleeding-associated factor (ebaf or LEFTY), the bone morphogenic proteins (BMPs), and the growth and differentiation factors (GDFs).120 In humans, TGF-β mRNA is distributed throughout the endometrium with lowest levels during in the proliferative phase and highest found in maternal decidua. There is evidence that TGF-β affects embryonic development, as mouse embryos exposed to TGF-β in vitro show a greater differentiation compared to those that were not exposed.140 Cultured human cytotrophoblasts secrete fetal fibronectin in the presence of TGF-β, likely promoting cellular adhesion. TGF-β may inhibit the production MMPs, thus preventing the normal menstrual breakdown of the endometrium.156 Mice with null-mutations for TGF-β have not been shown to have implantation failure, so the importance of TGF-β may be indirect.157
Other growth factors
Platelet-derived growth factors (PDGFs) are 30-kDa proteins that mediate intercellular signaling via tyrosine kinase receptors.120 PDGFs are composed of two subunits (A and B) and can result in one of three dimers (AA, AB, BB). The PDGFs stimulate proliferation and may have a role in embryonic development. The PDGF-α receptor null mutant mouse (the Patch mouse) has normal fertility and placentation, however, raising the question of whether PDGF is essential for implantation.158
Fibroblast growth factor (FGF) is an angiogenic growth factor that is highly expressed throughout the human menstrual cycle. FGF has been identified in four forms and the basic one (bFGF) increases endometrial stromal production of proteolytic enzymes and tissue plasminogen activator (t-PA), possibly aiding in trophoblast outgrowth and invasion.159 FGF-3 and FGF-4 and their receptors have been found in mouse embryos suggesting a role in preimplantation embryo development.160
Endometrial-embryonic dialogues
As evidenced by the information reviewed thus far, implantation is the result of an exceedingly complex series of cellular events (see Fig. 12). While much of the uterine preparation for implantation occurs without direct embryonic involvement, increasing evidence suggests that endometrium and the embryo may be targets for paracrine or juxtacrine signals that facilitate attachment and invasion. Cross-talk between the embryo and endometrium undoubtedly occurs. Data now exist that support a role for the endometrium in limiting placental invasion. Embryonic signals are also recognized as important for rescue of the CL that influences the reproductive tract directly through the action of progesterone. This early signal from the embryo may be essential for the recognition of pregnancy in primates and illustrates one example of this type of cross-talk.
As mentioned earlier, ECM may also play a role in embryo-endometrial signaling. Elaboration of the alternatively spliced form of fibronectin (fetal fibronectin) may form an adhesive interface between embryo and endometrium.161 Laminin produced by the blastocyst stimulates MMP-2 expression in both embryo and endometrium and the resulting proteolytic fragments of laminin have been shown to stimulate cell migration as an early inducer of the invasive phenotype.162 MMP production also promotes HB-EGF release from the endometrium and continues to promote embryo development.163, 164 Embryo quality is improved when it is cocultured with cells from the reproductive tract, likely as a result of these various interactions, and has led to improved outcomes in ART.165, 166
We compared the invasiveness of the embryo during implantation to the metastic spread of cancer cells.2 Complex cellular dialogues promote embryonic development and invasion, but also may at the same time prevent uncontrolled trophoblast invasiveness. One mechanism of regulation of embryonic invasiveness involves the TIMPs and inhibitors of plasmin. IGFBP-1 may also limit invasiveness through an interaction with the α5β1 integrin.155 We are just beginning to appreciate the enormous complexity of these interactions. A thorough understanding of these influential factors will likely lead to development of novel therapeutic means to influence the reproductive process. In the next section we explore some clinical examples of implantation failure and suggest strategies for managing these common problems.
CLINICAL EXAMPLES OF IMPLANTATION FAILURE
It is apparent from the data presented thus far that successful implantation depends on the synchronous development of both the endometrium and embryo. Loss of this synchrony results in the clinical manifestation of pregnancy loss and/or infertility in both animals and humans.167, 168 Luteal phase deficiency (LPD) is a major cause of infertility and recurrent pregnancy loss that results from either inadequate progesterone production or response.119 LPD has been difficult to study, however, because of a lack of consensus on its definition, and the estimated prevalence of the disorder ranges widely, from 4% to 60% of infertility patients. The true estimate likely will fall between 5% and 10% of truly infertile couples.119
Clinical features of luteal phase deficiency
The most stringent clinical diagnosis of LPD is based on a timed endometrial biopsy 7–9 days post-luteinizing hormone (LH) surge demonstrating a delay of 2 days in the dating criteria of Noyes and colleagues34 for histologic progression of endometrium. Other means of demonstrating LPD are basal body temperature (BBT) testing demonstrating less than 11 days of temperature elevation, low luteal phase serum progesterone levels (by either single or multiple assessments), or lack of production of a variety of markers of receptivity.27, 169, 170 Inadequate progesterone effect (from lack of either production or response) results in a histologic lag that delays the onset of uterine receptivity. The precise reason why this might result in reproductive failure is unclear, but may be a result of loss of embryo viability as it awaits a receptive endometrium. More likely, the fault is because of a failure of timely rescue of the CL. Inappropriate or untimely expression of many of the previously mentioned factors, such as PR, MUC-1, glycodelin (PP14), ebaf, MMPs, or the αvβ3 integrin and its ligand OPN, potentially could be used to identify hormonally inadequate cycles.
LPD is best viewed not as a single distinct entity, but rather as a final common result of various predisposing factors or causes. The causes of LPD are summarized in Table 2. A key point is that the functional capacity of the CL depends on the relative health of the preovulatory ovarian follicle from which it is derived. Therefore, as can be seen from Table 2, many factors that disrupt the hypothalamic-pituitary-ovarian axis, and thus result in fewer developed granulosa cells from which to form the CL, potentially lead to LPD.171
Table 2. Causes of luteal phase deficiency
Cause | Suspected Mechanism |
Medications | |
Opioids | Suppression of GnRH |
NSAIDs | Possible disruption of ovulation |
Clomiphene citrate | Antiestrogenic effect on uterus |
Phenothiazines | Elevated prolactin (inhibition of PIF) |
Endocrinopathy | |
Hyperprolactinemia | Altered GnRH release, direct ovarian effect |
Hyperandrogenism | Altered ovarian function |
Hypothyroidism | Elevated prolactin (increased TRH) |
Systemic disease | |
Eating disorders | Critical fat mass for ovulation |
Stress, excessive exercise or chronic illness | Suppression of GnRH (via endogenous opioids and CRH) |
Miscellaneous | |
Idiopathic | |
Endometrial abnormalities | Altered receptivity |
Inhibin abnormalities | Altered FSH release/folliculogenesis |
Early luteolysis | Direct or indirect CL effect |
Poor CL rescue | Delayed or inadequate implantation |
NSAIDs, nonsteroidal anti-inflammatory drugs; GnRH, gonadotropin-releasing hormone; PIF, prolactin inhibitory factor; TRH, thyrotropin-releasing hormone; CRH, corticotropic-releasing hormone; FSH, follicle stimulating hormone; CL, corpus luteum.
It is progesterone that causes the proliferative endometrium to undergo the characteristic secretory transformation and synthesis and secretion of factors essential for embryo implantation.172 Progesterone also influences the endometrial stroma, leading to the eventual conversion to the decidua, a highly specialized compartment that produces ECM, cytokines, and growth factors thought to be essential for implantation, but also to limit trophoblast invasion. It is noted that progesterone downregulates its receptor (PR) in the epithelium, but PR persists in the stroma.109, 113 This is a key event in development of the endometrium, marking a shift from epithelial to stromal responsiveness. Delay in PR (or ERα) downregulation may alter patterns of gene activation and thus render the endometrium unreceptive to embryo implantation.
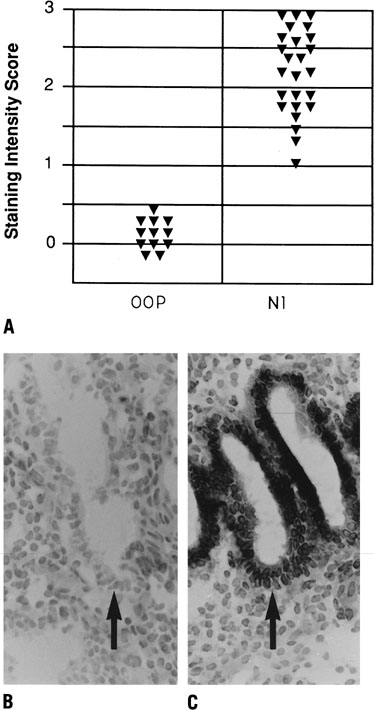
As mentioned previously, alterations in hormonal stimulation or response may result in endometrial defects. The first, the classically defined LPD, is referred to as a type I luteal deficiency and is characterized by a delay in the histologic progression of the luteal phase endometrium.119 This is a result of insufficient progesterone action on the endometrium. LPD is commonly found in women with unexplained infertility and recurrent pregnancy loss, but it is also commonly seen in these groups of women even when serum progesterone concentrations are not clearly low.173, 174 Balasch and Vanrell175 found that endometrial maturation was delayed by 2 or more days in 13.5% of infertile women and 32.5% of patients with a history of recurrent pregnancy loss, even though 86% overall had what they considered normal hormonal profiles. This demonstrates that LPD may result from factors influencing ovarian steroid production as well as the endometrium itself by means of its progesterone responsiveness. An important study by Wilcox and colleagues18 suggests that any delay in implantation will result in poor implantation efficiency. Women who had delayed implantation in this study were noted to have increasingly poor reproductive outcome and were reported to lack key proteins, such as αvβ3 integrins, that are normally present (Fig. 17).18
The utility of using histology to define LPD has, however, been seriously challenged by recent studies. First was a study in which the investigators attempted to refine the histologic criteria for dating first established by Noyes in 1950.176 They systematically evaluated 130 biopsies timed to specific days following detected LH surges, all done independently by three gynecologic pathologists, using 32 pre-defined histologic features to accurately predict the histologic date. The endometrial dating criteria were shown to be much less temporally distinct and discriminating than originally described with considerable intersubject, intrasubject, and interobserver variability. Neither traditional dating criteria nor any combination of the best performing histologic features could reliably distinguish any specific cycle day or narrow interval of days. Subsequent to this, the Reproductive Medicine Network published its findings from its multicenter endometrial biopsy study comparing fertile to infertile women with regards to endometrial dating and likelihood of having out-of-phase endometrium.177 They sampled 619 women and found that out-of-phase biopsies poorly discriminated fertile from infertile women, actually observing more out-of-phase biopsies in the fertile group (49%) than in the infertile group (43%). These findings combine to shed serious doubt in the clinical utility of endometrial biopsy in the assessment of infertile couples and in the role of endometrial histologic dating as a tool for research on uterine receptivity.
{kind=link}
Evidence may exist for a novel type of endometrial receptivity defect that is unrelated to endometrial histology. This concept was first developed from the study of women with endometriosis, and later in women with unexplained infertility,74 hydrosalpinges,76 and PCOS.80 We found that a subset of patients with endometriosis exhibited aberrant midluteal phase integrin expression despite normal histologic dating. Aberrant expression of various biomarkers has been described in such patients and may be caused by alterations in the signaling at the time of implantation. Such defects could be caused by the inhibitory effects of estrogen on epithelial expression of certain genes such as the αvβ3 integrin, interfering with the stimulatory effects of EGF or HB-EGF.118 Similar observations of integrin expression deficits were also made in patients with unexplained infertility74 and we recently extended these findings to include the effect of hydrosalpinges on endometrial function.76 Accumulating data support a detrimental effect on hydrosalpinx fluid on implantation in human reproduction, as an endometrial cavity bathed in hydrosalpinx fluid at the time of implantation decreases pregnancy rates in IVF.178, 179 Salpingectomy results in return of normal integrin expression in most women and yields improved pregnancy rates.76 Similar to endometriosis, inflammatory cytokines in the tubal fluid may alter normal endometrial function and account for this dysfunctional endometrium. This concept of two distinct types of uterine receptivity defects was previously depicted in Figure 15.
Treatment of luteal phase deficiency
Although early observational studies suggested improved cycle fecundity with treatment of LPD, subsequent trials have not been as convincing.169, 180 Because the causes of LPD are varied, directed treatment of an identified causative factor would be merited. As for an otherwise idiopathic LPD, several therapies have been used, none of which has been shown decidedly to be superior, and thus the choice of therapy is generally one of individual preference.119 It was noted in a study by Daly and coworkers181 that in a group of 29 women with idiopathic LPD (biopsy proven) who were randomly assigned to receive intravaginal progesterone, clomiphene citrate, or human menotropins, all groups achieved some pregnancies, but only in women whose treatment normalized their histologic dating.
EXOGENOUS PROGESTERONE
Because LPD results from inadequate progesterone activity on the endometrium, giving supplemental progesterone makes intuitive sense. Progesterone can be given by vaginal suppository, intramuscular injection, or micronized oral administration.119 Several observational studies have shown improved fecundity with progesterone administration; however, data from randomized clinical trials (RCT) are scarce.108, 175, 182, 183 Two studies randomly assigning women with LPD to either progesterone or clomiphene citrate showed no difference between the groups.182, 183 Progesterone administration in ART cycles results in a thicker endometrium and higher pregnancy rates.184 Because the granulosa is significantly disturbed by the retrieval process and is less adequately lutenized, it makes physiologic sense that progesterone supplementation would be beneficial. In general, however, the decision to give progesterone supplementation must consider the limited demonstrated benefit versus the theoretical risk of hyperandrogenizing a female infant through supraphysiologic uterine progesterone levels, although no congenital abnormalities have been directly attributable to progestin therapy.185
CLOMIPHENE CITRATE
Clomiphene citrate is the most frequently used drug for ovulation induction. Because LPD often results from a poor follicular phase, clomiphene citrate is also often used for LPD. When given to ovulatory women with LPD, the intent of clomiphene citrate treatment is to enhance follicular development and, in turn, the functional capacity of the resultant CL.108, 186 Paradoxically, Cook and colleagues187 reported that ovulatory women frequently develop LPD after receiving clomiphene citrate, an observation that suggests clomiphene citrate treatment may be detrimental in normally cycling women. Other reports show efficacy of clomiphene citrate on LPD, but only in women with profound degrees of histologic delay (5 days or more out of phase) or in women with demonstrated short luteal phase.188, 189 When given to women with unexplained infertility, clomiphene citrate reduces the incidence of histologic delay.190
HUMAN CHORIONIC GONADOTROPIN
Exogenous hCG stimulates the CL directly and represents an alternative to progesterone supplementation.119 Supplemental hCG is most appropriate when specific defects in preovulatory or postovulatory LH secretion can be identified or in the management of recurrent pregnancy loss when delayed implantation is suspected.184, 191 Recommendations for dosing vary widely, with some suggesting 5000–10,000 international units (IU) at the time of ovulation followed by 5000 IU 5–7 days later; alternatively 10,000 IU can be given in divided doses every 3 days after ovulation. Unfortunately, there are few studies evaluating the effectiveness of hCG.
KEY POINTS
- Implantation is a complex process involving ovarian steroids, a multitude of paracrine and autocrine factors, and interactions between multiple cell types.
- Timing of implantation is important and involves synchronous activities in the embryo, endometrium, and ovary.
- The mechanism of implantation is not precisely known but likely involves cell attachment receptors as well as specific ligands.
- Problems with implantation and the development of uterine receptivity may be common and involve disruptions in the signaling pathways involved in embryonic or endometrial receptivity.
- Histological dating has very limited utility in clinically distinguishing fertile from infertile women and as a research tool for the future
- New studies are needed to better understand receptivity defects and to devise better methods to treat infertility and for purposes of contraception.
REFERENCES
Navot D, Drews MR, Bergh P, et al: Age-related decline in female fertility is not due to diminished capacity of the uterus to sustain embryo implantation. Fertil Steril 61:97, 1994 |
|
Murray MJ, Lessey BA: Embryo implantation and tumor metastasis: Common pathways of invasion and angiogenesis. Semin Reprod Endocrinol 17:275, 1999 |
|
McLaren A, Michie D: Studies on the transfer of fertilized mouse eggs to uterine foster-mothers. J Exp Biol 33:394, 1974 |
|
Hodgen GD: Surrogate embryo transfer combined with estrogen-progesterone therapy in monkeys: Implantation, gestation, and delivery without ovaries. JAMA 250:2167, 1983 |
|
Psychoyos A: Uterine receptivity for nidation. Ann NY Acad Sci 476:36, 1986 |
|
Das S, Wang X, Paria B, et al: Heparin-binding EGF-like growth factor gene is induced in the mouse uterus temporally by the blastocyst solely at the site of its apposition: A possible ligand for interaction with blastocyst EGF-receptor in implantation. Development 120:1071, 1994 |
|
Chakraborty I, Das S, Wang J, et al: Developmental expression of the cyclo-oxygenase-1 and cyclo-oxygenase-2 genes in the peri-implantation mouse uterus and their differential regulation by the blastocyst and ovarian steroids. J Mol Endocrinol 16:107, 1996 |
|
Carson DD, Bagchi I, Dey SK, et al: Embryo implantation. Dev Biol 223:217, 2000 |
|
Parr M, Parr E: The implantation reaction. In Wynn RM, Jollie WP (eds): The Uterus. pp. 233, 277 New York, Plenum, 1989 |
|
Enders A, Schlafke S: Cytological aspects of trophoblast uterine interaction in early implantation. Am J Anat 125:1, 1969 |
|
Hoffman L, Olson G, Carson D, et al: Progesterone and implanting blastocysts regulate Muc1 expression in rabbit uterine epithelium. Endocrinology 139:266, 1998 |
|
Enders A, Schlafke S: Penetration of the uterine epithelium during implantation in the rabbit. Am J Anat 132:219, 1971 |
|
Kliman H, Nestler J, Sermasi E, et al: Purification characterization and in vitro differentiation of cytotrophoblasts from human term placentae. Endocrinology 118:1567, 1986 |
|
Coutifaris C, Babalola G, Feinberg R, et al: Purified human cytotropoblasts: Surrogates for the blastocyst in in vitro models of implantation? In Mashiach S (ed): Advances in Assisted Reproductive Technologies. pp. 687, 695 New York, Plenum Press, 1990 |
|
Bentin-Ley U, Horn T, Sjögren A, et al: Ultrastructure of human blastocyst-endometrial interactions in vitro. J Reprod Fertil 120:337, 2000 |
|
Hertig AT, Rock J, Adams ED: A description of 34 human ova within the first 17 days of development. Am J Anat 98:435, 1956 |
|
Yoshinaga K: Receptor concept in implantation research. In Yoshinaga K, Mori T (eds): Development of Preimplantation Embryos and their Environment. pp 379, 387 New York, Alan Liss, Inc., 1989 |
|
Wilcox AJ, Baird DD, Wenberg CR: Time of implantation of the conceptus and loss of pregnancy. N Engl J Med 340:1796, 1999 |
|
Bergh PA, Navot D: The impact of embryonic development and endometrial maturity on the timing of implantation. Fertil Steril 58:537, 1992 |
|
Buster J, Bustillo M, Rodi I, et al: Biologic and morphologic development of donated human ova recovered by nonsurgical uterine lavage. Am J Obstet Gynecol 153:211, 1985 |
|
O’Rahilly R, Muller F: Developmental Stages in Human Embryos, Publication 637. Washington, D.C., Carnegie Institution of Washington, 1987 |
|
Croxatto HB, Ortiz ME, Diaz S, et al: Studies on the duration of egg transport by the human oviduct. II. Ovum location at various intervals following luteinizing hormone peak Am J Obstet Gynecol 132:629, 1978 |
|
Mitchell MH, Mitchell MS, Swanson RJ, et al: Enhancement of in vitro murine embryo development by recombinant leukemia inhibitory factor. J Soc Gynecol Invest 1:215, 1994 |
|
Cullinan EB, Abbondanzo SJ, Anderson PS, et al: Leukemia inhibitory factor (LIF) and LIF receptor expression in human endometrium suggests a potential autocrine paracrine function in regulating embryo implantation. Proc Natl Acad Sci USA 93:3115, 1996 |
|
Martin KL, Barlow DH, Sargent IL: Heparin-binding epidermal growth factor significantly improves human blastocyst development and hatching in serum-free medium. Hum Reprod 13:1645, 1998 |
|
Fishel SB, Edwards RG, Evans CJ: Human chorionic gonadotropin secreted gy preimplantation embryos cultured in vitro. Science 223:816, 1984 |
|
Hay DL, Lopata A: Chorionic gonadotropin secretion in vitro. J Clin Endocrinol Metab 67:1322, 1988 |
|
Cohen J, Alikani M, Trowbridge J, et al: Implantation enhancement by selective assisted hatching using zona drillin of human embryos with poor prognosis. Hum Reprod 7:685, 1992 |
|
Cohen J, Wright G, Malter H, et al: Impairment of the hatching process following in vitro fertilization in the human and improvement of implantation by assisted hatching using micromanipulation. Hum Reprod 5:7, 1990 |
|
Stein A, Rufas O, Amit S, et al: Assisted hatching by partial zona dissection of human pre-embryos in patients with recurrent implantation failure after in vitro fertilization. Fertil Steril 63:838, 1995 |
|
Navot D, Bergh PA, Williams M, et al: An insight into early reproductive processes through the in vivo model of ovum donations. J Clin Endocrinol Metab 72:408, 1991 |
|
Gonen Y, Casper RF, Jacobson W, et al: Endometrial thickness and growth during ovarian stimulation: A possible predictor of implantation in in vitro fertilization. Fertil Steril 52:446, 1989 |
|
Abdalla HI, Brooks AA, Johnson MR, et al: Endometrial thickness: a predictor of implantation in ovum recipients? Hum Reprod 9:363, 1994 |
|
Noyes RW, Hertig AT, Rock J: Dating the endometrial biopsy. Fertil Steril 1:3, 1950 |
|
Martel D, Frydman R, Glissant M, et al: Scanning electron microscopy of postovulatory human endometrium in spontaneous cycles and cycles stimulated by hormone treatment. J Endocrinol 114:319, 1987 |
|
Martel D, Monier MN, Roche D, Psychoyos A: Hormonal dependence of pinopod formation at the uterine luminal surface. Hum Reprod 6:597, 1991 |
|
Nikas G, Drakas P, Loutradis D, et al: Uterin pinopods as markers of the ‘nidation window’ in cycling women receiving exogenous oestadiol and progesterone. Hum Reprod 10:1208, 1995 |
|
Savouret JF, Chauchereau A, Misrahi M, et al: The progesterone receptor. Biological effects of progestins and anti-progestins Hum Reprod 9S:7, 1994 |
|
Cowell TP: Implantation and development of mouse eggs transferred to the uterus of non-progestational mice. J Reprod Fertil 19:239, 1969 |
|
Fanchin R, Righini C, Olivennes F, et al: Uterine contractions at time of embryo transfer alter pregnancy rates after in vitro fertilization. Hum Reprod 13:1968, 1998 |
|
Lesny P, Killick SR, Tetlow RL, et al: Embryo transfer-can we learn anything from the observation of junctional zone contractions? Hum Reprod 13:1540, 1998 |
|
Aplin JD, Charlton AD, Ayad S: An immunohistochemical study of endometrial extracellular matrix during the menstrual cycle and first trimester of pregnancy. Cell Tissue Res 253:231, 1988 |
|
Bilalis DA, Klentzeris LD, Fleming S: Immunohistochemical localization of extracellular matrix proteins in luteal phase endometrium of fertile and infertile patients. Hum Reprod 11:2713, 1996 |
|
Hewitt K, Beer AE, Grinnell F: Disappearance of anionic sites from the surface of the rat endometrial epithelium at the time of blastocyst implantation. Biol Reprod 21:691, 1979 |
|
Murphy CR, Rogers AW: Effects of ovarian hormones on cell membranes in the rat uterus. III. The surface carbohydrates at the apex of the luminal epithelium Cell Biophys 3:305, 1981 |
|
Surveyor GA, Gendler SJ, Pemberton L, et al: Differential expression of Muc-1 at the apical cell surface of mouse uterine epithelial cells. FASEB J 7:1151a, 1993 |
|
Aplin JD, Hey NA: MUC1, endometruim and embryo implantation. Biochem Soc Trans 23:826, 1995 |
|
Woessner JF: Matrix metalloproteinases and their inhibitors in connective tissue remodeling. FASEB J 5:2145, 1991 |
|
Emonard H, Grimaud JA: Matrix metalloproteinases. A review Cell Mol Biol 36:131, 1990 |
|
Tabibzadeh S: The signals and molecular pathways involved in human menstruation, a unique process of tissue destruction and remodeling. Mol Hum Reprod 2:77, 1996 |
|
Salamonsen LA, Woolley DE: Matrix metalloproteinases in normal menstruation. Hum Reprod 11(Suppl 2):124, 1996 |
|
Nagase H: Activation mechanisms of matrix metalloproteinases. Biol Chem Hoppe-Seyler 378:151, 1997 |
|
Coussens LM, Werb Z: Matrix metalloproteinases and the development of cancer. Chem Biol 3:895, 1996 |
|
Gospodarowicz D, Greenburg G, Birdwell CR: Determination of cellular shape by the extracellular matrix and its correlation with the control of cellular growth. Cancer Res 38:4155, 1978 |
|
Werb Z, Tremble P, Damsky CH: Regulation of extracellular matrix degradation by cell-extracellular matrix interactions. Cell Diff Dev 32:299, 1990 |
|
Brooks PC, Stromblad S, Sanders LC, et al: Localization of matrix metalloproteinase MMP-2 to the surface of invasive cells by interaction with integrin αvβ3. Cell 85:683, 1996 |
|
Huppertz B, Kertschanska S, Demir AY, et al: Immunohistochemistry of matrix metalloproteinases (MMP), their substrates, and their inhibitors (TIMP) during trophoblast invasion in the human placenta. Cell Tissue Res 291:133, 1998 |
|
Librach CL, Werb Z, Fitzgerald ML, et al: 92-kD type IV collagenase mediates invasion of human cytotrophoblasts. J Cell Biol 113:437, 1991 |
|
Marzusch K, Ruck P, Dietl JA, et al: Immunohistochimical localization of tissue inhibitor of metalloproteinases-2 (TIMP-2) in first trimester human placental decidua. Eur J Obstet Gynecol Reprod Biol 68:105, 1996 |
|
Tabibzadeh S, Babaknia A: The signals and molecular pathways involved in adhesio and tissue invasion. Hum Reprod 10:1579, 1995 |
|
Fujimoto J, Hori M, Ichigo S, et al: Comparative study on expression of plasminogen activator inhibitor 1 and its mRNA in endometrial cancers and normal endometria. Tumor Biol 18:13, 1997 |
|
Khamsi F, Armstrong DT, Zhang XQ: Expression of urokinase-type plasminogen activator in human preimplantation embryos. Mol Hum Reprod 2:273, 1996 |
|
Queenan JT, Kao LC, Arboleda CE, et al: Regulation of urokinase-type plasminogen activator production by cultured human cytotrophoblasts. J Biol Chem 262:10903, 1987 |
|
Zini JM, Murray SC, Graham CH, et al: Characterization of urokinase receptor expression by human placental trophoblasts. Blood 79:2917, 1992 |
|
Axelrod HR: Altered trophoblast functions in implantation-defective mouse embryos. Dev Biol 108:185, 1985 |
|
Loskutoff DJ, Sawdey M, Mimuro J: Type I plasminogen activator. Prog Hemostasis Thromb 9:87, 1989 |
|
Glueck CJ: Plasminogen activator inhibitor activity: An independent risk factor for the high miscarriage rate during pregnancy in women with polycystic ovary syndrome. Metab Clin Exp 48:1589, 1999 |
|
Gris JC: Case control study of the frequency of thrombohilic disorders in couples with late foetal loss and no thrombotic antecednt (NOHA5). Thromb Haemost 81:891, 1999 |
|
Sutherland AE, Calarco PG, Damshky CH: Developmental regulation of integrin expression at the time of implantation in the mouse embryo. Development 119:1175, 1993 |
|
Damsky CH, Fitzgerald ML, Fisher SJ: Distribution patterns of extracellular matrix components and adhesion receptors are intricately modulated during first trimester cytotrophoblast differentiation along the invasive pathway, in vivo. J Clin Invest 89:210, 1992 |
|
Lessey BA, Damjanovich L, Coutifaris C, et al: Integrin adhesion molecules in the human endometrium. Correlation with the normal and abnormal menstrual cycle J Clin Invest 90:188, 1992 |
|
Lessey BA, Castelbaum AJ, Buck CA, et al: Further characterization of endometrial integrin during the menstrual cycle and in pregnancy. Fertil Steril 62:497, 1994 |
|
Tabibzadeh S: Patterns of expression of integrin molecules in human endometrium throughout the menstrual cycle. Hum Reprod 7:876, 1992 |
|
Lessey BA, Castelbaum AJ, Sawin SJ, et al: Integrins as markers of uterine receptivity in women with primary unexplained infertility. Fertil Steril 63:535, 1995 |
|
Lessey BA, Castelbaum AJ, Sawin SJ, et al: Aberrant integrin expression in the in endometrium of women with endometriosis. J Clin Enocriol Metab 79:643, 1994 |
|
Meyer WR, Castelbaum AJ, Harris JE, et al: Hydrosalpinges adversely affect markers of uterine receptivity. Hum Reprod 12:1393, 1997 |
|
Lessey BA, Ilesanmi AO, Sun J, et al: Luminal and glandular endometrial epithelium express integrins differentially throughout the menstrual cycle: Implication s for implantation, contraception, and infertility. Am J Reprod Immunol 35:195, 1996 |
|
Armant DR, Kaplan HA, Mover H, et al: The effect of hexapeptides on attachment and outgrowth of mouse blastocysts cultured in vitro: Evidence for the involvement of the cell recognition tripeptide Arg-Gly-Asp. Proc Natl Acad Sci USA 83:6751, 1986 |
|
Yelian FD, Yang Y, Hirata JD, et al: Molecular interactions between fibronectin and integrins during mouse blastocyst outgrowth. Mol Reprod Dev 41:435, 1995 |
|
Apparao KBC, Lovely LP, Gui Y, et al: Elevated endometrial androgen receptor expression in women with polycystic ovarian syndrome. Biol Reprod 66:297, 2002 |
|
Ilesanmi AO, Hawkins DA, Lessey BA: Immunohistochemical markers of uterine receptivity in the human endometrium. Microscopy Res Tech 25:208, 1993 |
|
Tulppala M, Julkunen M, Tiitinen A, et al: Habitual abortion is accompanied by low serum levels of placental protein 14 in the luteal phase of the fertile cycle. Fertil Steril 63:792, 1995 |
|
Fukuda MN, Sato T, Nakayama J, et al: Trophinin and tastin, a novel cell adhesion molecule complex with potential involvement in embryo implantation. Genes Dev 9:1199, 1995 |
|
Mac Calman CD, Furth EE, Omigbodun A, et al: Regulated expression of cadherin-11 in human epithelial cells: role for cadherin-11 in trophoblast-endometrium interactions? Dev Dyn 206:201, 1996 |
|
Albers A, Thie M, Hohn HP, et al: Differential expression and localization of integrins and CD44 in the membrane domains of human uterine epithelial cells during the menstrual cycle. Acta Anat 153:12, 1995 |
|
Stewart CL, Kaspar P, Brunet LJ, et al: Blastocyst implantation depends on maternal expression of leukmia inhibitory factor. Nature 359:76, 1992 |
|
Bhatt H, Brunet LJ, Stewart CL: Uterine expression of leukemia inhibitory factor coincides with the onset of blastocyst implantation. Proc Natl Acad Sci USA 88:11408, 1991 |
|
Das SK, Wang XN, Paria BC, et al: Heparin-binding EGF-like growth factor gene is induced in the mouse uterus temporally by the blastocyst solely at the site of apposition: A possible ligand for interaction with blastocyst EGF-receptor in implantation. Development 120:1071, 1994 |
|
Giudice LC: Endometrial growth factors and proteins. Semin Reprod Endocrinol 13:93, 1995 |
|
Apparao KBC, Murray MJ, Fritz MA, et al: Osteopontin and its receptor avb3 integrin are coexpressed in the human endometrium during the menstrual cycle but regulated differentially. J Clin Endocrinol Metab 86:4991, 2001 |
|
Ruoslahti E, Pierschbacher MD: New perspectives in cell adhesion: RDG and integrins. Science 238:491, 1987 |
|
Johnson GA, Spencer TE, Burghardt RC, et al: Progesterone modulation of osteopontin gene expression in the ovine uterus. Biol Reprod 62:1315, 2000 |
|
Lindenberg S, Hyttel P, Sjogren A, et al: A comparative study of attachment of human, bovine, and mouse blastocysts to uterine epithelial monolayer. Hum Reprod 4:446, 1989 |
|
Lindenberg S: Ultrastructure in human implantation: Transmission and scanning electron miroscopy. Bailieres Clin Obstet Gynaecol 5:1, 1991 |
|
Lindenberg S, Hyttel P, Lenz S, et al: Ultrastructure of the early human implantation in vitro. Hum Reprod 1:533, 1986 |
|
R. Alon and S. Feigelson, Semin. Immunol. 14, 93 (2002) |
|
O. Genbacev, et al, Science, 299(5605), 405 (2003) |
|
T. Lai, et al., Feril Steril, 85(3), 761 (2006) |
|
Werb Z: ECM and cell surface proteolysis: Regulating cellular ecology. Cell 91:493, 1997 |
|
Werb Z, Tremble PM, Behrendtsen O, et al: Signal transduction through the fibronectin receptor induces collagenase and stromelysin gene expression. J Cell Biol 109:877, 1989 |
|
Enders AC: Implantation (embryology). Encyclopedia Hum Biol 4:423, 1991 |
|
Brenner RM, West NB: Hormonal regulation of the reproductive tract in female mammals. Ann Rev Physiol 37:273, 1975 |
|
Baulieu EE: Contragestion and other clinical applications of RU-486, and antiprogesterone at the receptor. Science 245:1351, 1989 |
|
Matsuzaki S, Uehara S, Murakami T, et al: Quantitative analysis of estrogen receptor alpha and beta messenger ribonucleic acid levels in normal endometrium and ovarian endometriotic cysts using a real-time reverse transcription-polymerase chain reaction assay. Fertil Steril 74:753, 2000 |
|
Mote PA, Balleine RL, McGowan EM, et al: Colocalization of progesterone receptors A and B by dual immunofluorescent histochemistry in human endometrium during the menstrual cycle. J Clin Endocrinol Metab 84:2963, 1999 |
|
Bamberger AM, Milde-Langosch K, Loning T, et al: The glucocorticoid receptor is specifically expressed in the stromal compartment of the human endometrium. J Clin Endocrinol Metab 86:5071, 2001 |
|
Katzenellenbogen BS: Dynamics of steroid hormone recptor action. Annu Rev Physiol 42:17, 1980 |
|
Sanborn BM, Kuo KS, Held B: Estrogen and progestogen binding site concentrations in human endometrium and cervix throughout the menstrual cycle and in tissue from women taking oral contraceptives. J Steroid Biochem 9:951, 1978 |
|
Lessey BA, Killam AP, Metzger DA, et al: Immunohistochemical analysis of human uterine estrogen and progesterone receptors throughout the menstrual cycle. J Clin Endocrinol Metab 67:334, 1988 |
|
de Ziegler D, Cornel C, Bergeron C, et al: Controlled preparation of the endometrium with exogenous estradiol and progesterone in women having functioning ovaries. Fertil Steril 56:851, 1991 |
|
Lessey BA, Apparao KBC, Young SL, et al: Endometrial estrogen receptor-α expression is increased in women with defects inuterine receptivity: A unifying concept for implantation failure [abstract]? J Soc Gynecol Invest 9S:493, 2002 |
|
Garcia E, Bouchard P, De Brux J, et al: Use of immunoctyochemistry of progesterone and estrogen receptors for endometrialdating. J Clin Endocrinol Metab 67:80, 1988 |
|
Savouret JF, Chauchereau A, Misrahi M, et al: The progesterone receptor. Biological effects of progestins and anti-progestins Hum Reprod 9S:7, 1994 |
|
Somkuti SG, Yuan LW, Fritz MA, et al: Epidermal growth factor and sex steroids dynamically regulate a marker ofendometrial receptivity in Ishikawa cells. J Clin Endocrinol Metab 82:2192, 1997 |
|
Cunha GR, Bigsby RM, Cooke PS, et al: Stromal-epithelial interactions in adult organs: Review. Cell Differ 17:137, 1985 |
|
Kurita T, Young P, Brody JR, et al: Stromal progesterone receptors mediate the inhibitory effects of progesterone on estrogen-induced uterine epithelial cell deoxyribonucleic acid synthesis. Endocrinology 139:4708, 1998 |
|
Cooke PS, Buchanan DL, Young P, et al: Stromal estrogen receptors mediate mitogenic effects of estradiol on uterine epithelium. Proc Natl Acad Sci USA 94:6535, 1997 |
|
Lessey BA, Gui Y, Apparao KBC, et al: Heparin-binding EGF-like growth factor (HB-EGF) in the human endometrium: Apotential paracrine role during implantation. Mol Reprod Dev (in press). |
|
Fritz MA, Lessey BA: Defective luteal function. In Fraser IS, Jansen R, Lobo RA, Whitehead M (eds): Estrogens and Progestogens in Clinical Practice. pp 437, London, Churchill Livingstone, 1998 |
|
Kao LC, Giudice LC: Growth factors and implantation. In Lessey BA (ed): Infertility and Reproductive Medicine Clinics of North America: Implantation. p. 281, Philadelphia, WB Saunders Co., 2001 |
|
Clark SC, Kamen R: The human hematopoietic colony-stimulating factors. Science 236:1229, 1987 |
|
Pollard JW, Hunt JS, Wkitor-Jedrzejczak W, et al: A pregnancy defect in the osteopetrotic (op/op) mouse demonstrates the requirement for CSF-1 in female fertility. Dev Biol 148:273, 1991 |
|
Kauma SW, Aukerman SL, Eierman D, et al: Colon-stimulating factor-1 and c-fms expression in human endometrial tissues and placenta during the menstrual cycle and early pregnancy. J Clin Endocrinol Metab 73:746, 1991 |
|
Haimovici F, Anderson DJ: Effects of growth factors and growth factor-extracellular matrix interactions on mouse trophoblast outgrowth in vitro. Biol Reprod 49:124, 1993 |
|
Hilton DJ, Gough NM: Leukemia inhibitory factor: a biological perspective. J Cell Biochem 46:21, 1991 |
|
Bhatt H, Brunet LJ, Stewart CL: Uterine expression of leukemia inhibitory factor coincides with the onset of blastocyst implantation. Proc Natl Acad Sci USA 88:11408, 1991 |
|
Cullinan EB, Abbondanzo SJ, Anderson PS, et al: Leukemia inhibitory factor (LIF) and LIF receptor expression in human endometrium suggests a potential autocrine paracrine function in regulating embryo implantation. Proc Natl Acad Sci USA 93:3115, 1996 |
|
Nachtigall MJ, Kliman HJ, Feinberg RF, et al: The effect of leukemia inhibitory factor (LIF) on trophoblast differentiation: A potential role in human implantation. J Clin Endocrinol Metab 81:801, 1996 |
|
Dinaello CA: The biology of interleukin-1. Chem Immunol 51:1, 1992 |
|
Meisser A, Chardonnens D, Campana A, et al: Effects of tumour necrosis factor-alpha, interleukin-1 alpha, macrophage colony stimulating factor and transforming growth factor beta on trophoblastic matrix metalloproteinases. Mol Hum Reprod 5:252, 1999 |
|
Sheth KV, Roca GL, al-Sedairy ST, et al: Prediction of successful embryo implantation by measuring interleukin-1 alpha and immunosuppressive factor(s) in preimplantation embryo culture fluid. Fertil Steril 55:952, 1991 |
|
Frank GR, Brar AK, Jikihara H, et al: Interleukin-1β and the endometrium: An inhibitor of stromal cell differentiation and possible autoregulator of decidualization in humans. Biol Reprod 52:184, 1995 |
|
Simon C, Frances A, Piquette G, et al: Interleukin-1 system in the materno-trophoblast unit in human implantation: Immunohistochemical evidence for autocrine/paracrine function. J Clin Endocrinol Metab 78:847, 1994 |
|
De los Santos MJ, Mercader A, Frances A, et al: Role of endometrial factors in regulating secretion of components of the immunoreactive human embryonic interleukin-1 system during emmbryonic development. Biol Reprod 54:563, 1996 |
|
Simon C, Frances A, Lee BY, et al: Immunohistochemical localization, identificationa and regulation of the interleukin-1 receptor antagonist in the human endometrium. Hum Reprod 10:2472, 1995 |
|
Simon C, Frances A, Piquette GN, et al: Embryonic implantation mice is blocked by interleukin-1 receptor antagonist. Endocrinology 134:521, 1994 |
|
Abbondanzo SJ, Cullinan EB, McIntyre K, et al: Reproduction in mice lacking a functional type 1 IL-1 receptor. Endocrinology 137:3598, 1996 |
|
Fisher DA, Lakshmanan J: Metabolism and effects of epidermal growth factor and related growth factors in mammals. Endocr Rev 11:418, 1990 |
|
Beerli RR, Hunes NE: Epidermal growth factor-related peptides activate distinct subsets of ErbB receptors and differ in their biological activities. J Biol Chem 271:6071, 1996 |
|
Paria BC, Dey SK: Preimplantation embryo development in vitro: Cooperative interactions amongembryos and role of growth factors. Proc Natl Acad Sci USA 874756:1990 |
|
Machida T, Taga M, Minaguchi H: Effects of epidermal growth factor and transforming growth factor alpha on themouse trophoblast outgrowth in vitro. Eur J Endocrinol 133:741, 1995 |
|
Paria BC, Das SK, Andrews GK, et al: Expression of the epidermal growth factor gene is regulated in mouse blastocysts during delayed implantation. Proc Natl Acad Sci USA 90:55, 1993 |
|
Johnson DC, Chatterjee S: Embryo implantation in the rat uterus induced by epidermal growth factor. J Reprod Fertil 99:557, 1993 |
|
Das SK, Lim H, Wang J, et al: Inappropriate expression of human transforming growth factor (TGF)-α in the uterus of transgenic mouse causes downregulation of TGF-β receptors and delays the blastocyst-attachment reaction. J Mol Endocrinol 18:243, 1997 |
|
Hofmann GE, Scott RTJ, Bergh PA, et al: Immunohistochemical localization of epidermal growth factor in human endometrium, decidua, and placenta. J Clin Endocrinol Metab 73:882, 1991 |
|
Chegini N, Rossi MJ, Masterson BJ: Platelet-derived growth factor (PDGF), epidermal growth factor (EGF), and EGF and PDGF beta-receptors in human endometriual tissue-localization and in vitro action. Endocrinology 130:2373, 1992 |
|
Fazlebas AT, Hild-Petito S, Verhage HG: Secretory proteins and growth factos of the baboon (Papio anubis) uterus: Potential roles in pregnancy. Cell Biol Int 18:1145, 1994 |
|
Yoo HJ, Barlow DH, Bardon HJ: Temporal and spatial regulation of expression of heparin-binding epidermal growth factor-like growth factor in the human endometrium: A possible role in blastocyst implantation. Dev Genet 21:102, 1997 |
|
Wang XN, Das SK, Damm D, et al: Differential regulation of heparin-binding epidermal growth factor-like growth factor in the adult ovariectomized mouse uterus by progesterone and estrogen. Endocrinology 135:1264, 1994 |
|
Nonoshita LD, Wathen NC, Dsupin BA, et al: Insulin-like growth factors (IGF’s), IGF-binding proteins (IGFBP’s), and proteolyzed IGFBP-3 in embryonic cavities in early human pregnancy: Their potential relevance to maternal-embryonic and fetal interactions. J Clin Endocrinol Metab 79:1249, 1994 |
|
Zhou J, Dsupin BA, Giudice LC, et al: Insulin-like growth factor system gene expression in human endometrium during the menstrual cycle. J Clin Endocrinol Metab 79:1723, 1994 |
|
Guidice LC, Lamson G, Rosenfeld RG, et al: Insulin-like growth factor-II (IGF-II) and IGF binding proteins in human endometrium. Ann NY Acad Sci 626:295, 1991 |
|
Bell SC, Patel SR, Jackson JA, et al: Major secretory protein of human decidualized endometrium in pregnancy is an insulin-like growth factor-binding protein. J Endocrinol 118:317, 1988 |
|
Irwin JC, de las Fuentes L, Dsupin BA, et al: Insulin-like growth factor regulation of human endometrial stromal cell function: Coordinate effects on insulin-like growth factor binding protein-1, cell proliferation and prolactin secretion. Regul Pept 48:165, 1993 |
|
Jones JI, Gockerman A, Busby WH, et al: Insulin-like growth factor binding protein 1 stimulates cell migration and binds to the alpha 5/beta 1 integrin by means of its ARG-GLY-ASP sequence. Proc Natl Acad Sci USA 90:10553, 1993 |
|
Osteen KG, Bruner KL, Eisenburg E, et al: Progesterone induced transforming growth factor-β (TGF-β) suppressesendometrial metalloproteinases but TGF-β alone cannot block endometriotic-like lesions innude mice. O-071:S35. Presented at the Annual Meeting of the American Fertility Society 1995 |
|
Kallapur S, Ormsby I, Doetschaman T: Strain dependency of TGFβ1 function during embryogenesis. Mol Reprod Dev 52:341, 1999 |
|
Smith EA, Seldin MF, Martinez L: Mouse platelet-derived growth factor receptor alpha gene is deleted in W19H and patch mutations on chromosome 5. Proc Natl Acad Sci USA 88:4811, 1991 |
|
Nuttall RK, Kennedy TG: Epidermal growth factor and basic fibroblast growth facto increase the production of matrix metalloproteinases during in vitro decidulization of rat endometrial stromal cells. Endocrinology 141:629, 2000 |
|
Rappolee DA, Patel Y, Jacobson K: Expression of fibroblast growth facctor receptors in peri-implantation mouse embryos. Mol Reprod Dev 51:252, 1998 |
|
Feinberg RF, Kliman HJ, Lockwood CJ: Is oncofetal fibronectin a trophoblast glue for human implantation? Am J Pathol 138:537, 1991 |
|
Giannelli G, Faulk-Marziller J, Schiraldi O, et al: Induction of cell migration by matrix metalloproteinase-2 cleavage of laminin-5. Science 277:225, 1997 |
|
Suzuki M, Raab G, Moses MA, et al: Matrix metalloproteinase-3 releases active heparin-binding EGF-like growth factor by cleavage at a specific juxtamembrane site. J Biol Chem 272:31730, 1997 |
|
Martin KL, Barlow DH, Sargent IL: Heparin-binding epidermal growth factor significantly improves human blastocyst development and hatching in serum-free medium. Hum Reprod 13:1645, 1998 |
|
Anderson TL, Sieg SM, Hodgen GD: Membrane compostition of endometrial epithelium: Molecular markers of uterine receptivity to implantation. Hum Reprod 3:513, 1988 |
|
Gardner DK, Vella P, Lane M, et al: Culture and transfer of human blastocysts increases implantation rates and reduces the need for multiple embryo transfers. Fertil Steril 69:84, 1998 |
|
Pope WF: Uterine asynchrony: A cause of embryonic loss. Biol Reprod 39:999, 1988 |
|
Stephenson MD: Frequency of factors associated with habitual abortion in 197 couples. Fertil Steril 66:24, 1996 |
|
Soules MR, Wiebe RH, Aksel S, et al: The diagnosis and therapy of luteal phase deficiency. Fertil Steril 28:1033, 1977 |
|
Li TC, Lenton EA, Dockery P, et al: A comparison of some clinical and endocrinological features between cycles withnormal and defective luteal phases in women with unexplained infertility. Hum Reprod 5:805, 1990 |
|
Seif MW, Aplin JD, Buckley CH: Luteal phase defect: The possibility of an immunohistochemical diagnosis. Fertil Steril 51:273, 1989 |
|
Nakajima ST, Gibson M: Pathophysiology of luteal-phase deficiency in human reproduction. Clin Obstet Gynecol 34:167, 1991 |
|
Joshi SG: Progestin-regulated proteins of the human endometrium. Semin Reprod Endocrinol 1:221, 1983 |
|
McNeely MJ, Soules MR: The diagnosis of luteal phase deficiency: A critical review. Fertil Steril 50:1, 1988 |
|
Balasch J, Vanrell JA: Luteal phase deficiency: An inadequate endometrial response to normal hormone stimulation. Int J Fertil 31:368, 1986 |
|
M.Murray, et al, Fertil Steril, 81(5), 1333 (2004) |
|
C. Coutifaris, et al., Fertil Steril, 82(5), 1264 (2004) |
|
Andersen AN, Yue Z, Meng FJ, et al: Low implantation rate after in vitro fertilization in patients with hydrosalpinges diagnosed by ultrasonography. Hum Reprod 9:1935, 1994 |
|
Kassabji M, Sims JA, Butler L, et al: Reduced pregnancy outcome in patients with unilateral or bilateral hydrosalpinx after in vitro fertilization. Eur J Obstet Gynecol Reprod Biol 56:129, 1994 |
|
Karamardian LM, Grimes DA: Luteal phase deficiency: Effect of treatment on pregnancy rates. Am J Obstet Gynecol 167:1391, 1992 |
|
Daly DC: The endometrium and the luteal phase defect. Semin Reprod Endocrinol 1:237, 1983 |
|
Huang KE: The primary treatment of luteal phase inadequacy: Progesterone versus clomiphene citrate. Am J Obstet Gynecol 155:824, 1986 |
|
Murray DL, Reich L, Adashi EY: Oral clomiphene citrate and vaginal progesterone suppositories in the treatment of luteal phase dysfunction: A comparative study. Fertil Steril 51:35, 1989 |
|
Segal S, Casper RF: Progesterone supplementation increases luteal phase endometrial thickness and oestradiol levels in in vitro fertilization. Hum Reprod 7:1210, 1992 |
|
Scialli AR: Developmental effects of progesterone and its derivatives. Reprod Toxicol 2:3, 1988 |
|
Dodson KS, McNaughton MC, Coutts JRT: Infertility in women with apparently ovulatory cycles II. The effects of clomiphene treatment on the profiles of gonadotropins and sex steroid hormones in peripheral plasma Br J Obstet Gynaecol 82:625, 1975 |
|
Cook CL, Schroeder JA, Yussman MA, et al: Induction of luteal phase defect with clomiphene citrate. Am J Obstet Gynecol 149:613, 1984 |
|
Downs KA, Gibson M: Climphene citrate therapy for luteal phase defect. Fertil Steril 39:34, 1983 |
|
Quagliarello J, Weiss G: Clomiphene citrate in the management of infertility associated with shortened luteal phase. Fertil Steril 31:373, 1979 |
|
Rodin DA, Fisher AM, Clayton RN: Cycle abnormalities in infertile women with regular menstrual cycles: Effects of clomiphene citrate treatment. Fertil Steril 62:42, 1994 |
|
Jones GS, Aksel SS, Wentz AC: Serum progesterone values in the luteal phase defects. Obstet Gynecol 44:706, 1974 |